

-
摘要: 近年来, 智能体集群的能量高效利用(Energy efficient utilization, EEU)机制已经成为多智能体系统领域的热点问题, 如何使用有限的能量资源实现系统性能最优是该问题的核心研究内容. 考虑到智能体集群与生物族群的相似性, 探究生物族群的能量高效利用机制对提升智能体集群节能性能有着重要的研究价值. 为此, 首先介绍不同生物族群中蕴含的能量利用机制, 并根据节能方式的差异分成3类, 流体优势利用机制、流体阻碍克服机制和热量交换与扩散机制; 然后对这些机制进行总结与分析, 并提出一种具有一般性的能量高效利用模型; 最后, 探讨能量高效利用机制在多智能体系统应用中面临的挑战和发展趋势.Abstract: The energy efficient utilization (EEU) mechanism of agent clusters has become a hot topic in the multi-agent system field. The core research content of this topic is how to use limited energy resources to optimize multi-agent system performance. Considering the similarity between the agent clusters and the biological colonies, exploring the energy efficient utilization mechanism of biological colonies has important research value in improving the energy utilization performance of intelligent agent clusters. Firstly, this paper introduces the energy utilization mechanism of multiple biological colonies, and classifies them according to the differences in energy saving methods, fluid advantage utilization mechanism, fluid obstacle overcoming mechanism and heat exchange and diffusion mechanism. Then these mechanisms are summarized and analyzed, and a general model of efficient energy utilization is proposed. Finally, the challenges and development trends of energy efficient utilization mechanisms in multi-agent applications are discussed.1)
1 1 本文中流体是指生物族群长期生存的液体(海水)和气体(空气). -
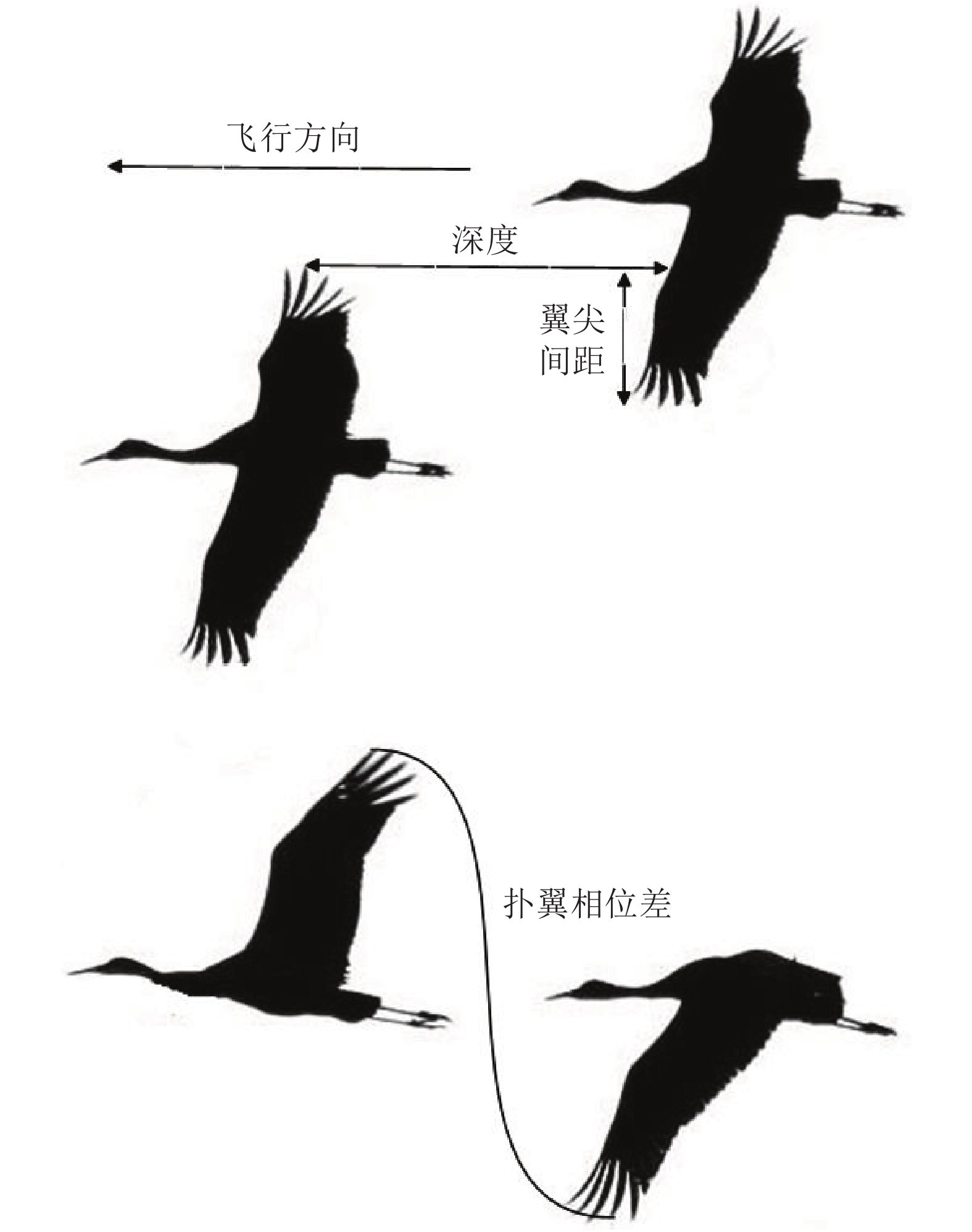
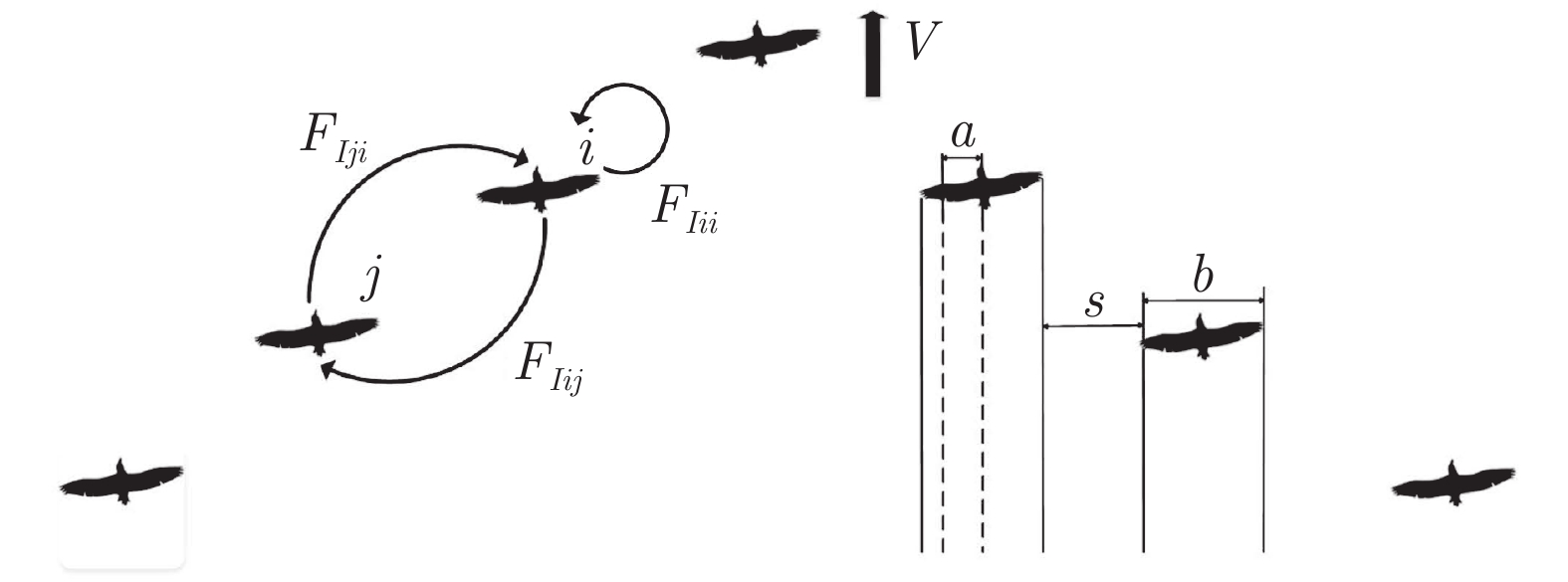
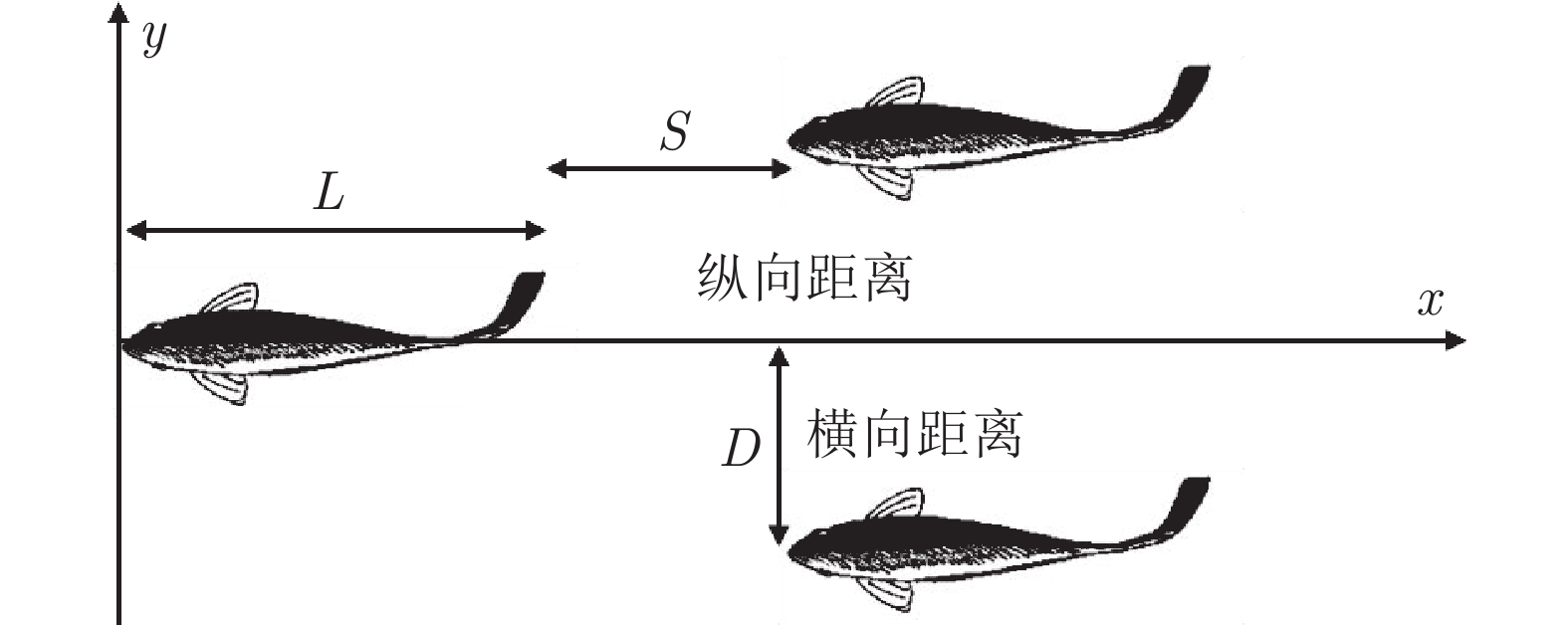
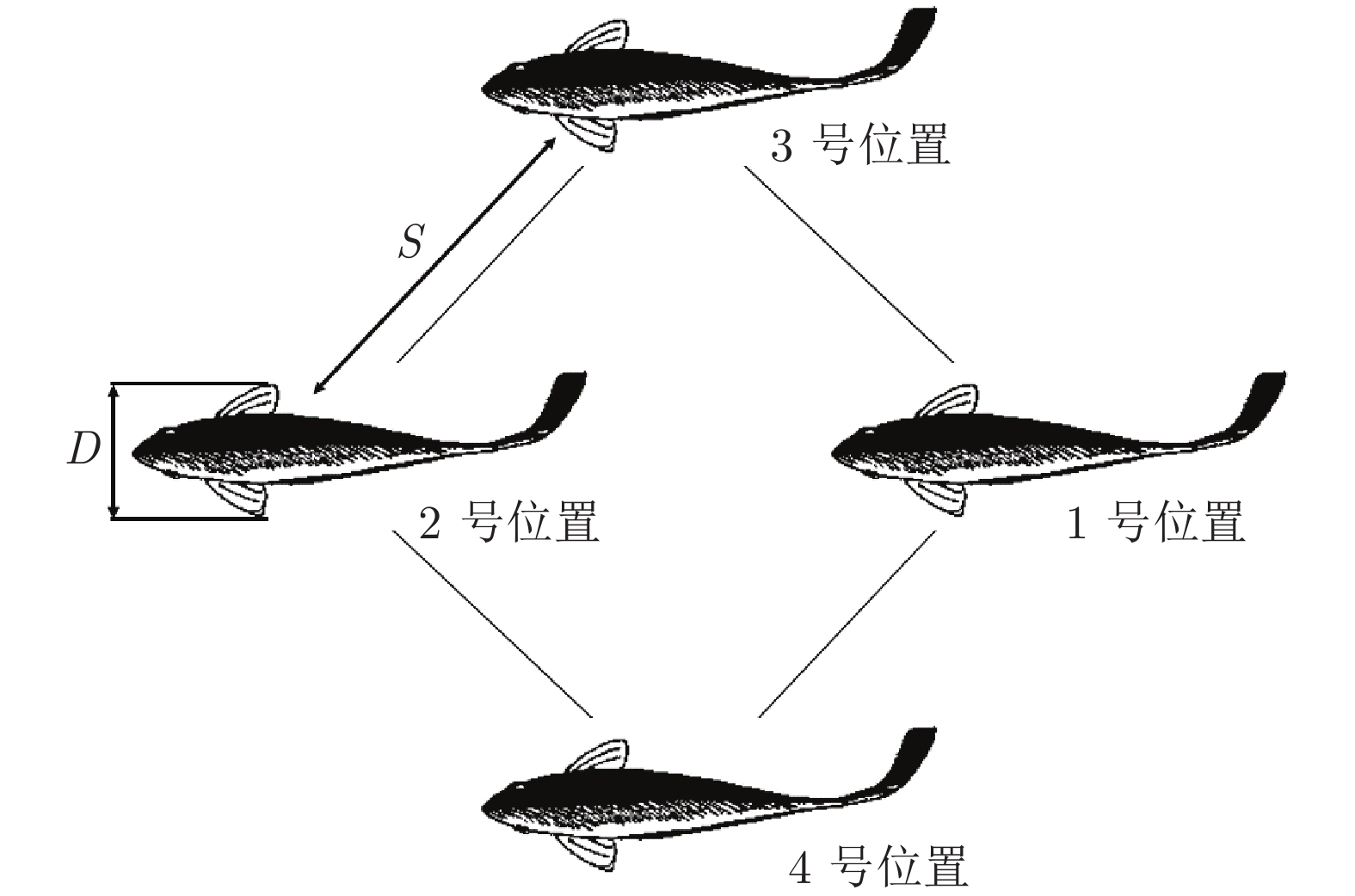
图 3 相邻鸟类间的“翼尖间距”、“深度”和“扑翼相位差”定义
Fig. 3 Definitions of “wing tip spacing”, “depth” and “flapping wing phase difference”
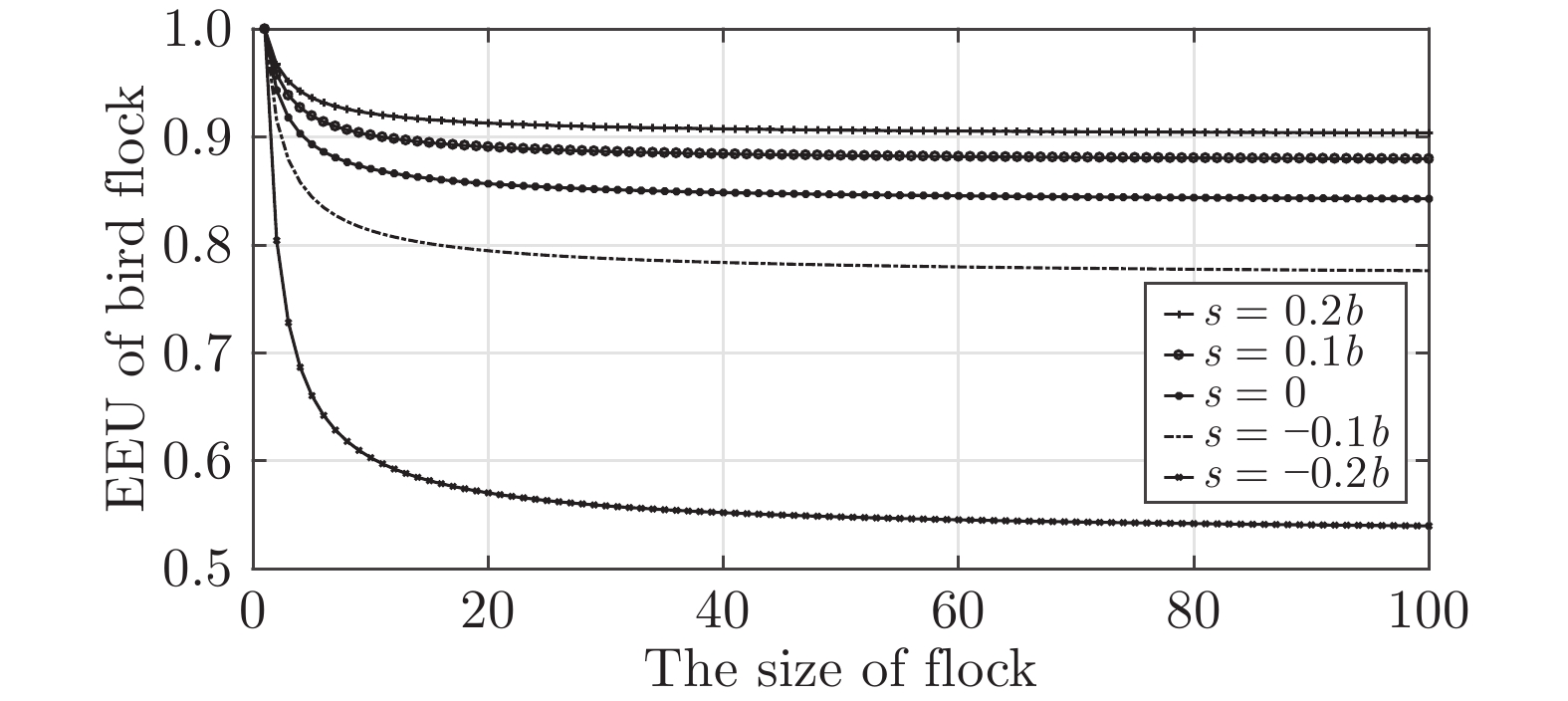
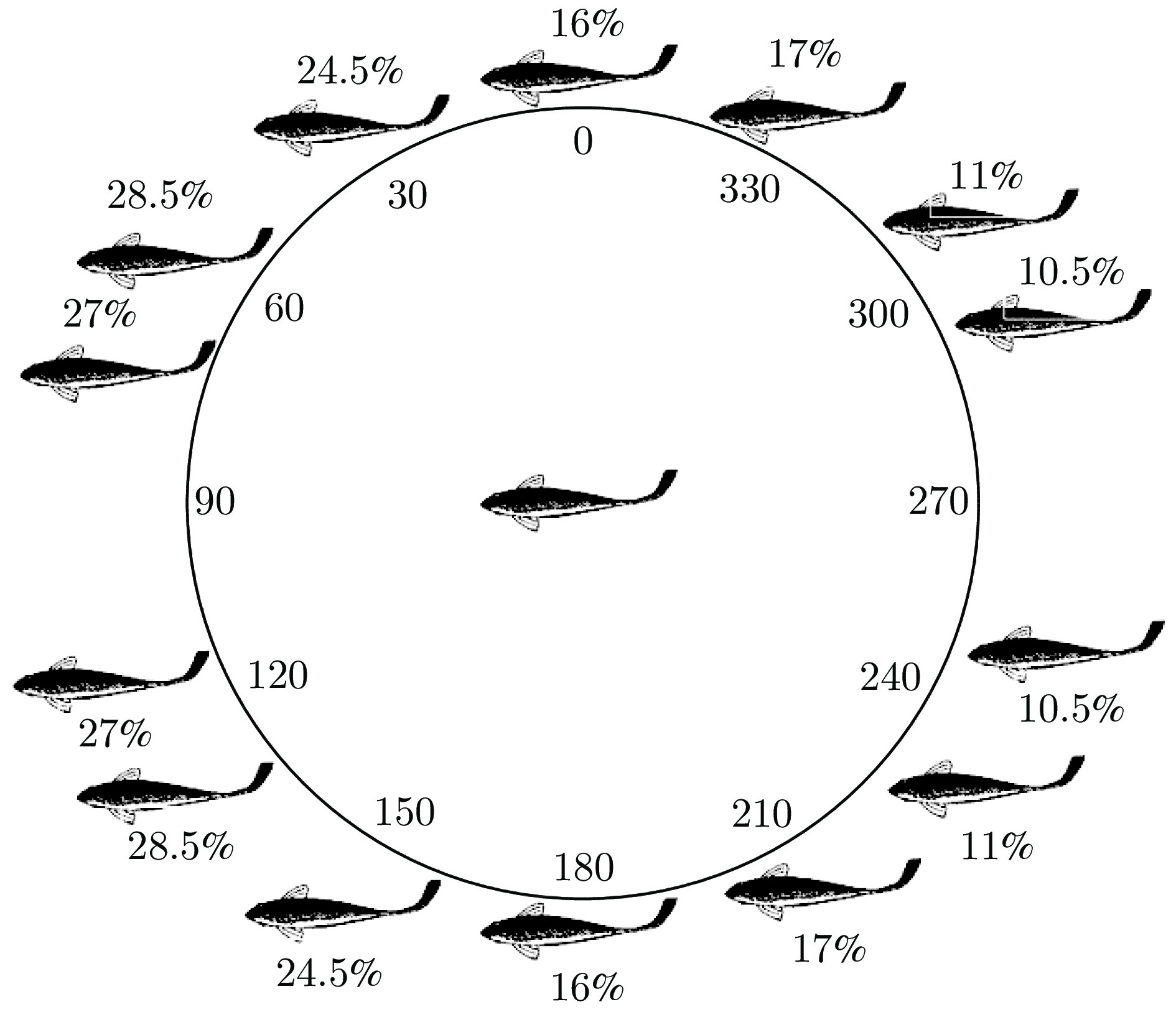
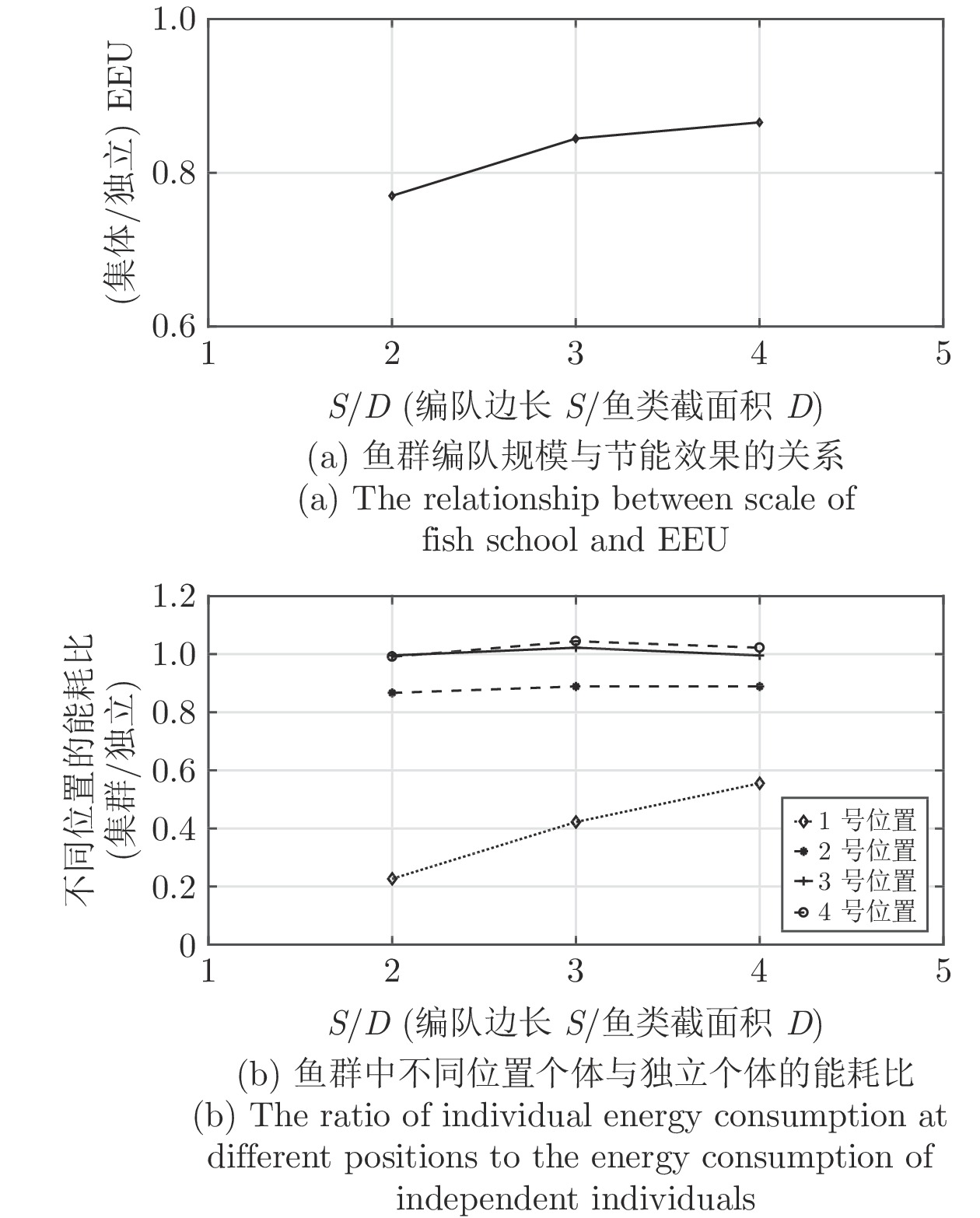
图 5 鸟群能量利用机制与集群规模$ n $和翼尖间距$ s $的关系
Fig. 5 Relationship between EEU of bird flock and the size $ n $ and wing tip spacing $ s $

图 12 南极磷虾集群 ((a)不同规模生物群体在聚集和分散情况下的能耗情况[104]; (b)磷虾运动时流体扰动的影响[108]; (c)磷虾群中不同的编队方式[109])
Fig. 12 Krill swarm ((a) Energy consumption of different group in non-swarming and swarming condition[104]; (b) Hydrodynamic disturbance from the motion of krill[108]; (c) Different formation method of krill swarm (Focal krill, FK)[109])

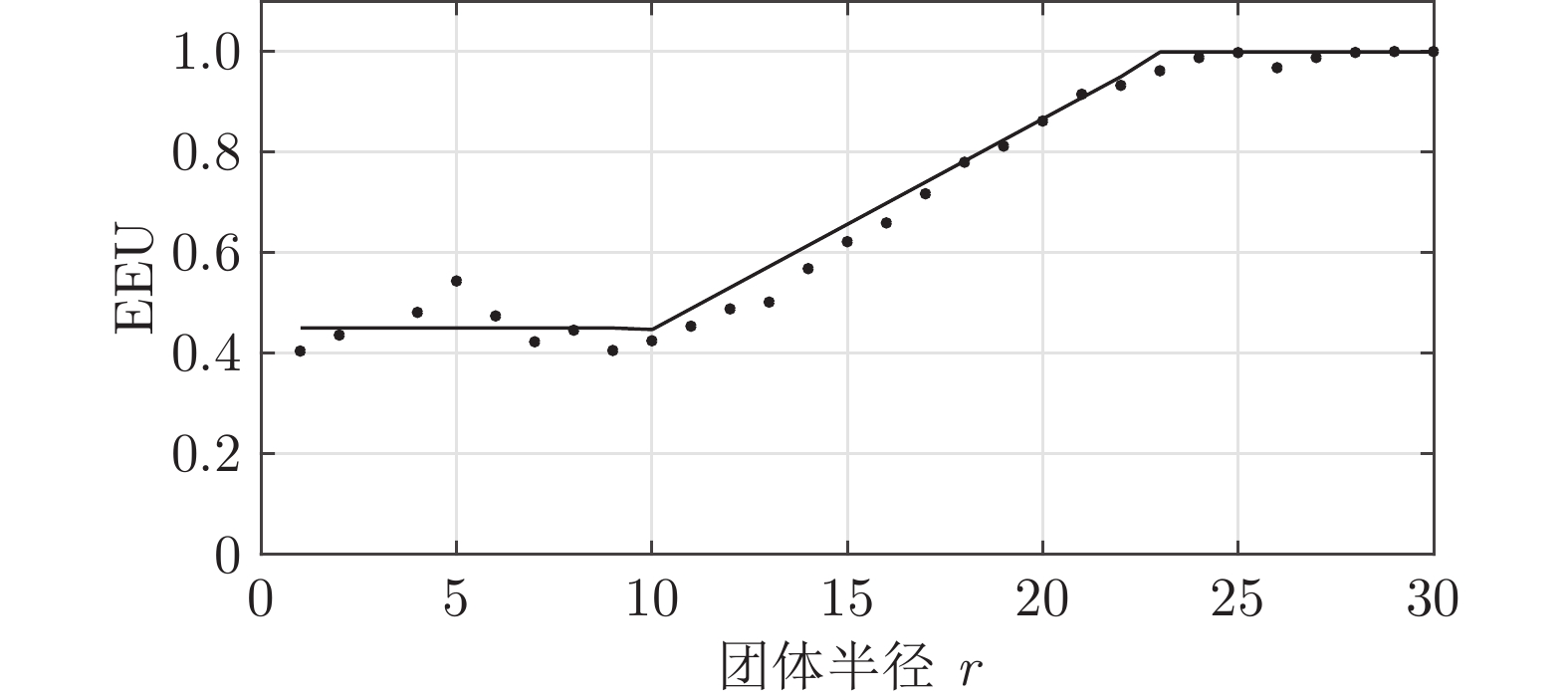
图 16 拥挤团体EEU随团体半径$ r $的变化趋势
Fig. 16 Relationship between EEU of huddling and radius $ r $
表 1 多圆柱体阻力表
Table 1 Drag coefficients of multi circle cylinders
位置序号 阻力系数 1 1.2158 2 0.4212 3 0.2191 4 0.1069 5 0.0861 6 0.0991  下载: 导出CSV
下载: 导出CSV
表 2 多种生物族群的能量高效利用机制总结
Table 2 Summary of energy efficient utilization mechanism in multiple biological clusters
族群种类 能量高效利用机制 实验数据 集群规模 EEU模型估计节能效果 参考文献 加拿大鹅 流体优势利用机制 能耗降低36.0% 55 9.4% ~ 45.3% (根据编队参数的差异) [57] 粉红足雁 流体优势利用机制 能耗降低14.0% 54 9.4% ~ 47.4% (根据编队参数的差异) [59] 白鹈鹕 流体优势利用机制 能耗降低11.4% ~ 14.0% 8 7.4% ~ 28.9% (根据编队参数的差异) [62] 鲭鱼 流体优势利用机制 摆动频率15.0% ~ 29.0% — 14.4% ~ 23.0% (根据编队间距的差异) [82] 海鲈鱼 流体优势利用机制 摆动频率9.0% ~ 14.0% 9 14.4% ~ 23.0% (根据编队间距的差异) [83] 欧洲拟鲤 流体优势利用机制 摆动频率7.3% ~ 11.6% 8 14.4% ~ 23.0% (根据编队间距的差异) [54] 鲻鱼 流体优势利用机制 摆动频率10.5% ~ 27.0% 8 14.4% ~ 23.0% (根据编队间距的差异) [87] 鳗鱼 流体优势利用机制 耗氧量30.0% 7 14.4% ~ 23.0% (根据编队间距的差异) [96] 南极磷虾 流体优势利用机制 耗氧量小7.2倍 — — [104] 棘刺龙虾 流体阻碍克服机制 65.0%阻力减免 19 70.6% (6只组成的队列) [117] 三叶虫 流体阻碍克服机制 — 3 30.6% (2只组成的队列) [129] 帝企鹅 热量交换与扩散机制 能耗降低51.0% — 最大节能效率不超过55.0% [138] 啮齿类动物幼崽 热量交换与扩散机制 — 100 最大节能效率不超过55.0% [148−149]
下载: 导出CSV
-
[1] Saad W, Bennis M, Mozaffari M, Lin X Q. Wireless Communications and Networking for Unmanned Aerial Vehicles. Cambridge: Cambridge University Press, 2020. [2] Ding J P, Mei H Y, Chih-Lin I, Zhang H, Liu W W. Frontier progress of unmanned aerial vehicles optical wireless technologies. Sensors, 2020, 20(19): Article No. 5476 doi: 10.3390/s20195476 [3] Baltaci A, Dinc E, Ozger M, Alabbasi A, Cavdar C, Schupke D. A survey of wireless networks for future aerial communications (FACOM). IEEE Communications Surveys & Tutorials, 2021, 23(4): 2833-2884 [4] Cai C W, Wu S, Jiang L Y, Zhang Z P, Yang S Y. A 500-W wireless charging system with lightweight pick-up for unmanned aerial vehicles. IEEE Transactions on Power Electronics, 2020, 35(8): 7721-7724 doi: 10.1109/TPEL.2020.2964023 [5] 李敏, 包富瑜, 王恒. 无人机使能的无线传感网总能耗优化方法. 自动化学报, DOI: 10.16383/j.aas.c220914Li Min, Bao Fu-Yu, Wang Heng. Optimization of total energy consumption for unmanned aerial vehicle-enabled wireless sensor networks. Acta Automatica Sinica, DOI: 10.16383/j.aas.c220914 [6] 罗小元, 杨帆, 李绍宝, 关新平. 多智能体系统的最优持久编队生成策略. 自动化学报, 2014, 40(7): 1311-1319Luo Xiao-Yuan, Yang Fan, Li Shao-Bao, Guan Xin-Ping. Generation of optimally persistent formation for multi-agent systems. Acta Automatica Sinica, 2014, 40(7): 1311-1319 [7] Shao L Y, Karci A E H, Tavernini D, Sorniotti A, Cheng M. Design approaches and control strategies for energy-efficient electric machines for electric vehicles—A review. IEEE Access, 2020, 8: 116900-116913 doi: 10.1109/ACCESS.2020.2993235 [8] Jiang B, Huang G S, Wang T, Gui J S, Zhu X Y. Trust based energy efficient data collection with unmanned aerial vehicle in edge network. Transactions on Emerging Telecommunications Technologies, 2022, 33(6): Article No. e3942 doi: 10.1002/ett.3942 [9] Boursianis A D, Papadopoulou M S, Diamantoulakis P, Liopa-Tsakalidi A, Barouchas P, Salahas G, et al. Internet of things (IoT) and agricultural unmanned aerial vehicles (UAVs) in smart farming: A comprehensive review. Internet of Things, 2022, 18: Article No. 100187 doi: 10.1016/j.iot.2020.100187 [10] Liu Y L, Dai H N, Wang Q B J, Shukla M K, Imran M. Unmanned aerial vehicle for internet of everything: Opportunities and challenges. Computer Communications, 2020, 155: 66-83 doi: 10.1016/j.comcom.2020.03.017 [11] Khan M T R, Muhammad Saad M, Ru Y, Seo J, Kim D. Aspects of unmanned aerial vehicles path planning: Overview and applications. International Journal of Communication Systems, 2021, 34(10): Article No. e4827 doi: 10.1002/dac.4827 [12] Chen J C, Ling F Y, Zhang Y, You T, Liu Y F, Du X Y. Coverage path planning of heterogeneous unmanned aerial vehicles based on ant colony system. Swarm and Evolutionary Computation, 2022, 69: Article No. 101005 doi: 10.1016/j.swevo.2021.101005 [13] Cho S W, Park H J, Lee H, Shim D H, Kim S Y. Coverage path planning for multiple unmanned aerial vehicles in maritime search and rescue operations. Computers & Industrial Engineering, 2021, 161: Article No. 107612 [14] Ahmed F, Mohanta J C, Keshari A, Yadav P S. Recent advances in unmanned aerial vehicles: A review. Arabian Journal for Science and Engineering, 2022, 47(7): 7963-7984 doi: 10.1007/s13369-022-06738-0 [15] Bouguettaya A, Zarzour H, Taberkit A M, Kechida A. A review on early wildfire detection from unmanned aerial vehicles using deep learning-based computer vision algorithms. Signal Processing, 2022, 190: Article No. 108309 doi: 10.1016/j.sigpro.2021.108309 [16] Saikin D A, Baca T, Gurtner M, Saska M. Wildfire fighting by unmanned aerial system exploiting its time-varying mass. IEEE Robotics and Automation Letters, 2020, 5(2): 2674-2681 doi: 10.1109/LRA.2020.2972827 [17] Neupane K, Baysal-Gurel F. Automatic identification and monitoring of plant diseases using unmanned aerial vehicles: A review. Remote Sensing, 2021, 13(19): Article No. 3841 doi: 10.3390/rs13193841 [18] Hu H M, Kaizu Y, Huang J J, Furuhashi K, Zhang H D, Li M, et al. Research on methods decreasing pesticide waste based on plant protection unmanned aerial vehicles: A review. Frontiers in Plant Science, 2022, 13: Article No. 811256 doi: 10.3389/fpls.2022.811256 [19] Jang G, Kim J, Yu J K, Kim H J, Kim Y, Kim D W, et al. Review: Cost-effective unmanned aerial vehicle (UAV) platform for field plant breeding application. Remote Sensing, 2020, 12(6): Article No. 998 doi: 10.3390/rs12060998 [20] McNelly B P, Whitcomb L L, Brusseau J P, Carr S S. Evaluating integration of autonomous underwater vehicles into port protection. In: Proceedings of the OCEANS, Hampton Roads. Hampton Roads, USA: IEEE, 2022. 1−8 [21] Al Abkal S, Talas R, Shaw S, Ellis T. The application of unmanned aerial vehicles in managing port and border security in the US and Kuwait: Reflections on best practice for the UK. International Journal of Maritime Crime & Security (IJMCS), 2020, 1(1): 26-33 [22] Gorodetsky V, Skobelev P, Marik V. System engineering view on multi-agent technology for industrial applications: Barriers and prospects. Cybernetics and Physics, 2020, 9(1): 13-30 [23] 杜永浩, 邢立宁, 蔡昭权. 无人飞行器集群智能调度技术综述. 自动化学报, 2020, 46(2): 222-241Du Yong-Hao, Xing Li-Ning, Cai Zhao-Quan. Survey on intelligent scheduling technologies for unmanned flying craft clusters. Acta Automatica Sinica, 2020, 46(2): 222-241 [24] Calegari R, Ciatto G, Mascardi V, Omicini A. Logic-based technologies for multi-agent systems: A systematic literature review. Autonomous Agents and Multi-Agent Systems, 2021, 35(1): Article No. 1 doi: 10.1007/s10458-020-09478-3 [25] Wieselsberger C. Beitrag zur Erklarung des Winkelfluges eineger Zugvogel. Motorluftschiffahrt, 1914, 3(5): 225-229 [26] Gould L L, Heppner F. The vee formation of Canada geese. The Auk, 1974, 165(11): 494-506 [27] Lissaman P B S, Shollenberger C A. Formation flight of birds. Science, 1970, 168(3934): 1003-1005 doi: 10.1126/science.168.3934.1003 [28] Weihs D. Hydromechanics of fish schooling. Nature, 1973, 241(5387): 290-291 doi: 10.1038/241290a0 [29] Belyayev V V. Zuyev G V. Hydrodynamic hypothesis of school formation in fishes. Problems of Ichthyology, 1969, 9(27): 578-584 [30] Fish F E. Energetics of swimming and flying in formation. Comments on Theoretical Biology, 1999, 5(35): 283-304 [31] Le Maho Y. The Emperor Penguin: A Strategy to Live and Breed in the Cold: Morphology, physiology, ecology, and behavior distinguish the polar emperor penguin from other penguin species, particularly from its close relative, the king penguin. American Scientist, 1977, 65(6): 680-693 [32] Ancel A, Visser H, Handrich Y, Masman D, Le Maho Y. Energy saving in huddling penguins. Nature, 1997, 385(6614): 304-305 doi: 10.1038/385304a0 [33] Herrnkind W. Queuing behavior of spiny lobsters. Science, 1969, 164(3886): 1425-1427 doi: 10.1126/science.164.3886.1425 [34] Chatterton B D E, Fortey R A. Linear clusters of articulated trilobites from Lower Ordovician (Arenig) strata at Bini Tinzoulin, North of Zagora, southern Morocco. Advances in Trilobite Research. Cuadernos del Museo Geominero, 2008, 24(9): 73-77 [35] Li W, Wang G G, Gandomi A H. A survey of learning-based intelligent optimization algorithms. Archives of Computational Methods in Engineering, 2021, 28(5): 3781-3799 doi: 10.1007/s11831-021-09562-1 [36] 刘成汉, 何庆. 融合多策略的黄金正弦黑猩猩优化算法. 自动化学报, 2023, 49(11): 2360−2373Liu Cheng-Han, He Qing. Golden sine chimp optimization algorithm integrating multiple strategies. ACTA AUTOMATICA SINICA, 2023, 49(11): 2360−2373 [37] 李雅丽, 王淑琴, 陈倩茹, 王小钢. 若干新型群智能优化算法的对比研究. 计算机工程与应用, 2020, 56(22): 1-12Li Ya-Li, Wang Shu-Qin, Chen Qian-Ru, Wang Xiao-Gang. Comparative study of several new swarm intelligence optimization algorithms. Computer Engineering and Applications, 2020, 56(22): 1-12 [38] 杨旭, 王锐, 张涛. 面向无人机集群路径规划的智能优化算法综述. 控制理论与应用, 2020, 37(11): 2291-2302Yang Xu, Wang Rui, Zhang Tao. Review of unmanned aerial vehicle swarm path planning based on intelligent optimization. Control Theory & Applications, 2020, 37(11): 2291-2302 [39] Qiu H X, Duan H B. A multi-objective pigeon-inspired optimization approach to UAV distributed flocking among obstacles. Information Sciences, 2020, 509: 515-529 doi: 10.1016/j.ins.2018.06.061 [40] Li L, Nagy M, Graving J M, Bak-Coleman J, Xie G M, Couzin I D. Vortex phase matching as a strategy for schooling in robots and in fish. Nature Communications, 2020, 11(1): Article No. 5408 doi: 10.1038/s41467-020-19086-0 [41] Wu X Y, He W, Wang Q, Meng T T, He X Y, Fu Q. A long-endurance flapping-wing robot based on mass distribution and energy consumption method. IEEE Transactions on Industrial Electronics, 2023, 70(8): 8215-8224 doi: 10.1109/TIE.2022.3213905 [42] Fu Q, Wang X Q, Zou Y, He W. A miniature video stabilization system for flapping-wing aerial vehicles. Guidance, Navigation and Control, 2022, 2(1): Article No. 2250001 doi: 10.1142/S2737480722500017 [43] Huang H F, He W, Fu Q, He X Y, Sun C Y. A bio-inspired flapping-wing robot with cambered wings and its application in autonomous airdrop. IEEE/CAA Journal of Automatica Sinica, 2022, 9(12): 2138-2150 doi: 10.1109/JAS.2022.106040 [44] Huang H F, He W, Wang J B, Zhang L, Fu Q. An all servo-driven bird-like flapping-wing aerial robot capable of autonomous flight. IEEE/ASME Transactions on Mechatronics, 2022, 27(6): 5484-5494 [45] He W, Mu X X, Zhang L, Zou Y. Modeling and trajectory tracking control for flapping-wing micro aerial vehicles. IEEE/CAA Journal of Automatica Sinica, 2021, 8(1): 148-156 doi: 10.1109/JAS.2020.1003417 [46] Pennycuick C J, Alerstam T, Hedenström A. A new low-turbulence wind tunnel for bird flight experiments at Lund University, Sweden. Journal of Experimental Biology, 1997, 200(10): 1441-1449 doi: 10.1242/jeb.200.10.1441 [47] Sachs G, Moelyadi M A. Effect of slotted wing tips on yawing moment characteristics. Journal of Theoretical Biology, 2006, 239(1): 93-100 doi: 10.1016/j.jtbi.2005.07.016 [48] Kawaguchi S, King R, Meijers R, Osborn J E, Swadling K M, Ritz D A, et al. An experimental aquarium for observing the schooling behaviour of Antarctic krill (Euphausia superba). Deep Sea Research Part II: Topical Studies in Oceanography, 2010, 57(7-8): 683-692 doi: 10.1016/j.dsr2.2009.10.017 [49] Hamner W M, Hamner P P. Behavior of Antarctic krill (Euphausia superba): Schooling, foraging, and antipredatory behavior. Canadian Journal of Fisheries and Aquatic Sciences, 2000, 57(S3): 192-202 doi: 10.1139/f00-195 [50] Catton K B, Webster D R, Kawaguchi S, Yen J. The hydrodynamic disturbances of two species of krill: Implications for aggregation structure. Journal of Experimental Biology, 2011, 214(11): 1845-1856 doi: 10.1242/jeb.050997 [51] Lopez U, Gautrais J, Couzin I D, Theraulaz G. From behavioural analyses to models of collective motion in fish schools. Interface Focus, 2012, 2(6): 693-707 doi: 10.1098/rsfs.2012.0033 [52] Viscido S V, Parrish J K, Grünbaum D. Individual behavior and emergent properties of fish schools: A comparison of observation and theory. Marine Ecology Progress Series, 2004, 273: 239-249 doi: 10.3354/meps273239 [53] Tian G Z, Zhang Y S, Feng X M, Hu Y S. Focus on bioinspired textured surfaces toward fluid drag reduction: Recent progresses and challenges. Advanced Engineering Materials, 2022, 24(1): Article No. 2100696 doi: 10.1002/adem.202100696 [54] Svendsen J C, Skov J, Bildsoe M, Steffensen J F. Intra-school positional preference and reduced tail beat frequency in trailing positions in schooling roach under experimental conditions. Journal of Fish Biology, 2003, 62(4): 834-846 doi: 10.1046/j.1095-8649.2003.00068.x [55] Wiese K, Ebina Y. The propulsion jet of Euphausia superba (Antarctic Krill) as a potential communication signal among conspecifics. Journal of the Marine Biological Association of the United Kingdom, 1995, 75(1): 43-54 doi: 10.1017/S0025315400015186 [56] Berger M. Formationsflug ohne Phasenbeziehung der Flügelschläge. Journal für Ornithologie, 1972, 113(2): 161-169 [57] Hainsworth F R. Precision and dynamics of positioning by Canada geese flying in formation. Journal of Experimental Biology, 1987, 128(1): 445-462 doi: 10.1242/jeb.128.1.445 [58] Bajec I L, Heppner F H. Organized flight in birds. Animal Behaviour, 2009, 78(4): 777−789 [59] Cutts C J, Speakman J R. Energy savings in formation flight of pink-footed geese. Journal of Experimental Biology, 1994, 189(1): 251-261 doi: 10.1242/jeb.189.1.251 [60] Badgerow J P, Hainsworth F R. Energy savings through formation flight? A re-examination of the vee formation. Journal of Theoretical Biology, 1981, 93(1): 41-52 doi: 10.1016/0022-5193(81)90055-2 [61] Ward S, Bishop C M, Woakes A J, Butler P J. Heart rate and the rate of oxygen consumption of flying and walking barnacle geese (Branta leucopsis) and bar-headed geese (Anser indicus). Journal of Experimental Biology, 2002, 205(21): 3347-3356 doi: 10.1242/jeb.205.21.3347 [62] Weimerskirch H, Martin J, Clerquin Y, Alexandre P, Jiraskova S. Energy saving in flight formation. Nature, 2001, 413(6857): 697-698 doi: 10.1038/35099670 [63] Bairlein F, Fritz J, Scope A, Schwendenwein I, Stanclova G, van Dijk G, et al. Energy expenditure and metabolic changes of free-flying migrating northern bald ibis. PLoS One, 2015, 10(9): Article No. e0134433 doi: 10.1371/journal.pone.0134433 [64] Nathan R, Spiegel O, Fortmann-Roe S, Harel R, Wikelski M, Getz W M. Using tri-axial acceleration data to identify behavioral modes of free-ranging animals: General concepts and tools illustrated for griffon vultures. Journal of Experimental Biology, 2012, 215(6): 986-996 doi: 10.1242/jeb.058602 [65] Späni D, Arras M, König B, Rülicke T. Higher heart rate of laboratory mice housed individually vs in pairs. Laboratory Animals, 2003, 37(1): 54-62 doi: 10.1258/002367703762226692 [66] Maeng J S, Park J H, Jang S M, Han S Y. A modeling approach to energy savings of flying Canada geese using computational fluid dynamics. Journal of Theoretical Biology, 2013, 320: 76-85 doi: 10.1016/j.jtbi.2012.11.032 [67] Kshatriya M, Blake R W. Theoretical model of the optimum flock size of birds flying in formation. Journal of Theoretical Biology, 1992, 157(2): 135-174 doi: 10.1016/S0022-5193(05)80618-6 [68] Mirzaeinia A, Hassanalian M, Lee K, Mirzaeinia M. Energy conservation of V-shaped swarming fixed-wing drones through position reconfiguration. Aerospace Science and Technology, 2019, 94: Article No. 105398 doi: 10.1016/j.ast.2019.105398 [69] Nudds R L, Rayner J M V. Scaling of body frontal area and body width in birds. Journal of Morphology, 2006, 267(3): 341-346 doi: 10.1002/jmor.10409 [70] Pennycuick C J, Fast P L F, Ballerstädt N, Rattenborg N. The effect of an external transmitter on the drag coefficient of a bird’s body, and hence on migration range, and energy reserves after migration. Journal of Ornithology, 2012, 153(3): 633-644 doi: 10.1007/s10336-011-0781-3 [71] Cutts C J, Metcalfe N B, Taylor A C. Aggression and growth depression in juvenile atlantic salmon: The consequences of individual variation in standard metabolic rate. Journal of Fish Biology, 1998, 52(5): 1026-1037 [72] Andersson M, Wallander J. Kin selection and reciprocity in flight formation?. Behavioral Ecology, 2004, 15(1): 158-162 doi: 10.1093/beheco/arg109 [73] Higdon J J L, Corrsin S. Induced drag of a bird flock. The American Naturalist, 1978, 112(986): 727-744 doi: 10.1086/283314 [74] Usherwood J R, Stavrou M, Lowe J C, Roskilly K, Wilson A M. Flying in a flock comes at a cost in pigeons. Nature, 2011, 474(7352): 494-497 doi: 10.1038/nature10164 [75] 张天栋, 王睿, 程龙, 王宇, 王硕. 鱼集群游动的节能机理研究综述. 自动化学报, 2021, 47(3): 475-488Zhang Tian-Dong, Wang Rui, Cheng Long, Wang Yu, Wang Shuo. Research on energy-saving mechanism of fish schooling: A review. Acta Automatica Sinica, 2021, 47(3): 475-488 [76] Spedding G. The cost of flight in flocks. Nature, 2011, 474(7352): 458-459 doi: 10.1038/474458a [77] Breder C M. Ortices and fish schools. Zoologica New York, 1965, 50(16): 97-114 [78] Weihs D. Some hydrodynamical aspects of fish schooling. Swimming and Flying in Nature. New York: Springer, 1975. 703−718 [79] Partridge B L, Pitcher T J. Evidence against a hydrodynamic function for fish schools. Nature, 1979, 279(5712): 418-419 doi: 10.1038/279418a0 [80] Blake R W. Fish Locomotion. Cambridge: Cambridge University Press, 1983. [81] Abrahams M V, Colgan P W. Fish schools and their hydrodynamic function: A reanalysis. Environmental Biology of Fishes, 1987, 20(1): 79-80 doi: 10.1007/BF00002028 [82] Zuyev G V. Belyayev V V. An experimental study of the swimming of fish in groups as examplified by the horsemackerel[Trachurus mediterraneus ponticus Aleev]. Journal of Ichthyology, 1970, 10(21): 545-549 [83] Herskin J, Steffensen J F. Energy savings in sea bass swimming in a school: Measurements of tail beat frequency and oxygen consumption at different swimming speeds. Journal of Fish Biology, 1998, 53(2): 366-376 doi: 10.1111/j.1095-8649.1998.tb00986.x [84] Gerstner C L. Use of substratum ripples for flow refuging by Atlantic cod, Gadus morhua. Environmental Biology of Fishes, 1998, 51(4): 455-460 doi: 10.1023/A:1007449630601 [85] Webb P W. The effect of solid and porous channel walls on steady swimming of steelhead trout Oncorhynchus mykiss. Journal of Experimental Biology, 1993, 178(1): 97-108 doi: 10.1242/jeb.178.1.97 [86] Liao J C. A review of fish swimming mechanics and behaviour in altered flows. Philosophical Transactions of the Royal Society B: Biological Sciences, 2007, 362(1487): 1973-1993 doi: 10.1098/rstb.2007.2082 [87] Marras S, Killen S S, Lindström J, McKenzie D J, Steffensen J F, Domenici P. Fish swimming in schools save energy regardless of their spatial position. Behavioral Ecology and Sociobiology, 2015, 69(2): 219-226 doi: 10.1007/s00265-014-1834-4 [88] Hemelrijk C, Reid D, Hildenbrandt H, Padding J. The increased efficiency of fish swimming in a school. Fish and Fisheries, 2015, 16(3): 511-521 doi: 10.1111/faf.12072 [89] Dai L Z, He G W, Zhang X, Zhang X. Stable formations of self-propelled fish-like swimmers induced by hydrodynamic interactions. Journal of the Royal Society Interface, 2018, 15(147): Article No. 20180490 doi: 10.1098/rsif.2018.0490 [90] Chen S Y, Fei Y H J, Chen Y C, Chi K J, Yang J T. The swimming patterns and energy-saving mechanism revealed from three fish in a school. Ocean Engineering, 2016, 122: 22-31 doi: 10.1016/j.oceaneng.2016.06.018 [91] Tesch F W, Thorpe J E. The Eel. Oxford: Blackwell Science Ltd, 2003. [92] Burgerhout E, Manabe R, Brittijn S A, Aoyama J, Tsukamoto K, van den Thillart G E E J M. Dramatic effect of pop-up satellite tags on eel swimming. Naturwissenschaften, 2011, 98(7): 631-634 doi: 10.1007/s00114-011-0805-0 [93] Palstra A, van Ginneken V, van den Thillart G. Cost of transport and optimal swimming speed in farmed and wild European silver eels (Anguilla anguilla). Comparative Biochemistry and Physiology Part A: Molecular & Integrative Physiology, 2008, 151(1): 37-44 [94] Sébert P, Scaion D, Belhomme M. High hydrostatic pressure improves the swimming efficiency of European migrating silver eel. Respiratory Physiology & Neurobiology, 2009, 165(1): 112-114 [95] van Ginneken V, Antonissen E, Müller U K, Booms R, Eding E, Verreth J, et al. Eel migration to the Sargasso: Remarkably high swimming efficiency and low energy costs. Journal of Experimental Biology, 2005, 208(7): 1329-1335 doi: 10.1242/jeb.01524 [96] Burgerhout E, Tudorache C, Brittijn S A, Palstra A P, Dirks R P, van den Thillart G E E J M. Schooling reduces energy consumption in swimming male European eels, Anguilla anguilla L. Journal of Experimental Marine Biology and Ecology, 2013, 448: 66-71 doi: 10.1016/j.jembe.2013.05.015 [97] 周梦园, 吴君钦, 夏乐, 黄敏. 智能仿生鱼系统的设计与实现. 计算机工程与设计, 2022, 43(5): 1467-1476Zhou Meng-Yuan, Wu Jun-Qin, Xia Le, Huang Min. Design and implementation of intelligent bionic fish system. Computer Engineering and Design, 2022, 43(5): 1467-1476 [98] 秦伟伟, 张建, 左新龙, 唐文献, 王子航. 基于锦鲤BCF摆动推进特性分析. 科学技术与工程, 2023, 23(8): 3200-3206Qin Wei-Wei, Zhang Jian, Zuo Xin-Long, Tang Wen-Xian, Wang Zi-Hang. Based on the analysis of the oscillating propulsion characteristics of koi BCF. Science Technology and Engineering, 2023, 23(8): 3200-3206 [99] 李广浩, 冯娜, 刘贵杰. 基于仿生鱼体结构的平板减阻方法. 水下无人系统学报, 2021, 29(1): 80-87Li Guang-Hao, Feng Na, Liu Gui-Jie. Flat drag reduction method based on biomimetic fish-body structure. Journal of Unmanned Undersea Systems, 2021, 29(1): 80-87 [100] 郝天泽, 肖华平, 刘书海, 张超, 马豪. 集成化智能软体机器人研究进展. 浙江大学学报(工学版), 2021, 55(2): 229-243Hao Tian-Ze, Xiao Hua-Ping, Liu Shu-Hai, Zhang Chao, Ma Hao. Research status of integrated intelligent soft robots. Journal of Zhejiang University (Engineering Science), 2021, 55(2): 229-243 [101] 教柳, 张保成, 张开升, 赵波. 两关节压力驱动柔性仿生机器鱼的设计与仿真. 力学学报, 2020, 52(3): 817-827Jiao Liu, Zhang Bao-Cheng, Zhang Kai-Sheng, Zhao Bo. Design and simulation of two-joint pressure-driven soft bionic fish. Chinese Journal of Theoretical and Applied Mechanics, 2020, 52(3): 817-827 [102] Borazjani I, Sotiropoulos F. Numerical investigation of the hydrodynamics of carangiform swimming in the transitional and inertial flow regimes. Journal of Experimental Biology, 2008, 211(10): 1541-1558 doi: 10.1242/jeb.015644 [103] Gonçalves R T, Hirabayashi S, Suzuki H. Experimental study on flow around an array of four circular cylinders. In: Proceedings of the Techno-Ocean (Techno-Ocean). Kobe, Japan: IEEE, 2016. 660−667 [104] Ritz D A. Is social aggregation in aquatic crustaceans a strategy to conserve energy? Canadian Journal of Fisheries and Aquatic Sciences, 2000, 57(S3): 59-67 doi: 10.1139/f00-170 [105] Patria M P, Wiese K. Swimming in formation in krill (Euphausiacea), a hypothesis: Dynamics of the flow field, properties of antennular sensor systems and a sensory-motor link. Journal of Plankton Research, 2004, 26(11): 1315-1325 doi: 10.1093/plankt/fbh122 [106] Hamner W M. Aspects of schooling in Euphausia superba. Journal of Crustacean Biology, 1984, 4(5): 67-74 doi: 10.1163/1937240X84X00507 [107] Kanda K, Takagi K, Seki Y. Movement of the larger swarms of Antarctic krill Euphausia superba population off enderby land during 1976-1977 season. Journal of the Tokyo University of Fisheries, 1982, 68(1-2): 25-45 [108] Yen J, Brown J, Webster D R. Analysis of the flow field of the krill, Euphausia pacifica. Marine and Freshwater Behaviour and Physiology, 2003, 36(4): 307-319 doi: 10.1080/10236240310001614439 [109] Murphy D W, Olsen D, Kanagawa M, King R, Kawaguchi S, Osborn J, et al. The three dimensional spatial structure of Antarctic krill schools in the laboratory. Scientific Reports, 2019, 9(1): Article No. 381 doi: 10.1038/s41598-018-37379-9 [110] Burns A L, Schaerf T M, Lizier J, Kawaguchi S, Cox M, King R, et al. Self-organization and information transfer in Antarctic krill swarms. Proceedings of the Royal Society B: Biological Sciences, 2022, 289(1969): Article No. 20212361 doi: 10.1098/rspb.2021.2361 [111] Tarling G A, Fielding S. Swarming and behaviour in Antarctic krill. Biology and Ecology of Antarctic Krill. Cham: Springer, 2016. 279−319 [112] Terushkin M, Fridman E. Network-based deployment of nonlinear multi agents over open curves: A PDE approach. Automatica, 2021, 129: Article No. 109697 doi: 10.1016/j.automatica.2021.109697 [113] Khan M W, Wang J. Multi-agents based optimal energy scheduling technique for electric vehicles aggregator in microgrids. International Journal of Electrical Power & Energy Systems, 2022, 134: Article No. 107346 [114] Herrnkind W F, Childress M J, Lavalli K L. Cooperative defence and other benefits among exposed spiny lobsters: Inferences from group size and behaviour. Marine and Freshwater Research, 2001, 52(8): 1113-1124 doi: 10.1071/MF01044 [115] Heeremans O, Rubie E, King M, Oviedo-Trespalacios O. Group cycling safety behaviours: A systematic review. Transportation Research Part F: Traffic Psychology and Behaviour, 2022, 91: 26-44 doi: 10.1016/j.trf.2022.09.013 [116] Pérez-Zuriaga A M, Moll S, López G, García A. Driver behavior when overtaking cyclists riding in different group configurations on two-lane rural roads. International Journal of Environmental Research and Public Health, 2021, 18(23): Article No. 12797 doi: 10.3390/ijerph182312797 [117] Bill R G, Herrnkind W F. Drag reduction by formation movement in spiny lobsters. Science, 1976, 193(4258): 1146-1148 doi: 10.1126/science.193.4258.1146 [118] Kanciruk P, Herrnkind W. Mass migration of spiny lobster, Panulirus argus (Crustacea: Palinuridae): Behavior and environmental correlates. Bulletin of Marine Science, 1978, 28(4): 601-623 [119] Herrnkind W F. Evolution and mechanisms of mass single-file migration in spiny lobster: Synopsis. Contributions in Marine Science, 1985, 27(S1): 197-211 [120] Herrnkind W F. Spiny lobsters: Patterns of movement. Biology and management of lobsters. Physiology and behavior, 1980, 1(12): 349-407 [121] Briceño F A, Polymeropoulos E T, Fitzgibbon Q P, Dambacher J M, Pecl G T. Changes in metabolic rate of spiny lobster under predation risk. Marine Ecology Progress Series, 2018, 598: 71-84 doi: 10.3354/meps12644 [122] Voelkl B, Portugal S J, Unsöld M, Usherwood J R, Wilson A M, Fritz J. Matching times of leading and following suggest cooperation through direct reciprocity during V-formation flight in ibis. Proceedings of the National Academy of Sciences of the United States of America, 2015, 112(7): 2115-2120 [123] Waters A, Blanchette F, Kim A D. Modeling huddling penguins. PLoS One, 2012, 7(11): Article No. e50277 doi: 10.1371/journal.pone.0050277 [124] Corrales-García A, Esteve J, Zhao Y L, Yang X L. Synchronized moulting behaviour in trilobites from the Cambrian Series 2 of South China. Scientific Reports, 2020, 10(1): Article No. 14099 doi: 10.1038/s41598-020-70883-5 [125] Mángano M G, Buatois L A, Waisfeld B G, Muñoz D F, Vaccari N E, Astini R A. Were all trilobites fully marine? Trilobite expansion into brackish water during the early Palaeozoic. Proceedings of the Royal Society B: Biological Sciences, 2021, 288(1944): Article No. 20202263 doi: 10.1098/rspb.2020.2263 [126] Bicknell R D C, Holmes J D, García-Bellido D C, Paterson J R. Malformed individuals of the trilobite Estaingia bilobata from the Cambrian Emu Bay Shale and their palaeobiological implications. Geological Magazine, 2023, 160(4): 803-812 doi: 10.1017/S0016756822001261 [127] Hou X G, Siveter D J, Aldridge R J, Siveter D J. Collective behavior in an early Cambrian arthropod. Science, 2008, 322(5899): 224-224 doi: 10.1126/science.1162794 [128] Radwański A, Kin A, Radwańska U. Queues of blind phacopid trilobites trimerocephalus: A case of frozen behaviour of early Famennian age from the Holy Cross Mountains, Central Poland. Acta Geologica Polonica, 2009, 59(4): 459-481 [129] Kin A, Błażejowski B. A new Trimerocephalus species (Trilobita, Phacopidae) from the late Devonian (early Famennian) of Poland. Zootaxa, 2013, 3626(3): 345-355 doi: 10.11646/zootaxa.3626.3.3 [130] Gutiérrez-Marco J C, Sá A A, García-Bellido D C, Rábano I, Valério M. Giant trilobites and trilobite clusters from the Ordovician of Portugal. Geology, 2009, 37(5): 443-446 doi: 10.1130/G25513A.1 [131] Fornarelli F, Oresta P, Lippolis A. Flow patterns and heat transfer around six in-line circular cylinders at low Reynolds number. JP Journal of Heat and Mass Transfer, 2015, 11(1): 1-28 doi: 10.17654/JPHMTFeb2015_001_028 [132] Gilbert C, Blanc S, Le Maho Y, Ancel A. Energy saving processes in huddling emperor penguins: From experiments to theory. Journal of Experimental Biology, 2008, 211(1): 1-8 doi: 10.1242/jeb.005785 [133] Gilbert C, Le Maho Y, Perret M, Ancel A. Body temperature changes induced by huddling in breeding male emperor penguins. American Journal of Physiology-Regulatory, Integrative and Comparative Physiology, 2007, 292(1): R176-R185 doi: 10.1152/ajpregu.00912.2005 [134] Ancel A, Gilbert C, Poulin N, Beaulieu M, Thierry B. New insights into the huddling dynamics of emperor penguins. Animal Behaviour, 2015, 110: 91-98 doi: 10.1016/j.anbehav.2015.09.019 [135] Alberts J R. Huddling by rat pups: Group behavioral mechanisms of temperature regulation and energy conservation. Journal of Comparative and Physiological Psychology, 1978, 92(2): 231-245 doi: 10.1037/h0077459 [136] Hayes J P, Speakman J R, Racey P A. The contributions of local heating and reducing exposed surface area to the energetic benefits of huddling by short-tailed field voles (Microtus agrestis). Physiological Zoology, 1992, 65(4): 742-762 doi: 10.1086/physzool.65.4.30158537 [137] Haim A, Van Aarde R J, Skinner J D. Burrowing and huddling in newborn porcupine: The effect on thermoregulation. Physiology & Behavior, 1992, 52(2): 247-250 [138] Gilbert C, McCafferty D, Le Maho Y, Martrette J M, Giroud S, Blanc S, et al. One for all and all for one: The energetic benefits of huddling in endotherms. Biological Reviews, 2010, 85(3): 545-569 doi: 10.1111/j.1469-185X.2009.00115.x [139] Gilbert C, Robertson G, Le Maho Y, Naito Y, Ancel A. Huddling behavior in emperor penguins: Dynamics of huddling. Physiology & Behavior, 2006, 88(4-5): 479-488 [140] Gilbert C, Robertson G, Le Maho Y, Ancel A. How do weather conditions affect the huddling behaviour of emperor penguins? Polar Biology, 2008, 31(2): 163-169 [141] Ancel A, Beaulieu M, Le Maho Y, Gilbert C. Emperor penguin mates: Keeping together in the crowd. Proceedings of the Royal Society B: Biological Sciences, 2009, 276(1665): 2163-2169 doi: 10.1098/rspb.2009.0140 [142] Zitterbart D P, Wienecke B, Butler J P, Fabry B. Coordinated movements prevent jamming in an emperor penguin huddle. PLoS One, 2011, 6(6): Article No. e20260 doi: 10.1371/journal.pone.0020260 [143] Alberts J R. Huddling by rat pups: Multisensory control of contact behavior. Journal of Comparative and Physiological Psychology, 1978, 92(2): 220-230 doi: 10.1037/h0077458 [144] Canals M, Bozinovic F. Huddling behavior as critical phase transition triggered by low temperatures. Complexity, 2011, 17(1): 35-43 doi: 10.1002/cplx.20370 [145] Conklin P, Heggeness F W. Maturation of tempeature homeostasis in the rat. American Journal of Physiology-Legacy Content, 1971, 220(2): 333-336 doi: 10.1152/ajplegacy.1971.220.2.333 [146] Malik S S, Fewell J E. Thermoregulation in rats during early postnatal maturation: Importance of nitric oxide. American Journal of Physiology-Regulatory, Integrative and Comparative Physiology, 2003, 285(6): R1366-R1372 doi: 10.1152/ajpregu.00280.2003 [147] Richter C P. Animal behavior and internal drives. The Quarterly Review of Biology, 1927, 2(3): 307-343 doi: 10.1086/394279 [148] Sealander Jr J A. The relationship of nest protection and huddling to survival of Peromyscus at low temperature. Ecology, 1952, 33(1): 63-71 doi: 10.2307/1931252 [149] Glancy J, Groß R, Stone J V, Wilson S P. A self-organising model of thermoregulatory huddling. PLoS Computational Biology, 2015, 11(9): Article No. e1004283 doi: 10.1371/journal.pcbi.1004283 [150] Glancy J, Groß R, Wilson S P. A minimal model of the phase transition into thermoregulatory huddling. In: Proceedings of the 2nd International Conference on Biomimetic and Biohybrid Systems. London, UK: Springer, 2013. 381−383 [151] 贺威, 丁施强, 孙长银. 扑翼飞行器的建模与控制研究进展. 自动化学报, 2017, 43(5): 685-696He Wei, Ding Shi-Qiang, Sun Chang-Yin. Research progress on modeling and control of flapping-wing air vehicles. Acta Automatica Sinica, 2017, 43(5): 685-696 [152] 尹曌, 贺威, 邹尧, 穆新星, 孙长银. 基于“雁阵效应”的扑翼飞行机器人高效集群编队研究. 自动化学报, 2021, 47(6): 1355-1367Yin Zhao, He Wei, Zou Yao, Mu Xin-Xing, Sun Chang-Yin. Efficient formation of flapping-wing aerial vehicles based on wild geese queue effect. Acta Automatica Sinica, 2021, 47(6): 1355-1367 [153] 付强, 张祥, 赵民, 张春华, 贺威. 仿生扑翼飞行器风洞实验研究进展. 工程科学学报, 2022, 44(4): 767-779Fu Qiang, Zhang Xiang, Zhao Min, Zhang Chun-Hua, He Wei. Research progress on the wind tunnel experiment of a bionic flapping-wing aerial vehicle. Chinese Journal of Engineering, 2022, 44(4): 767-779 [154] 汪婷婷, 何修宇, 邹尧, 付强, 贺威. 面向扑翼飞行机器人的飞行控制研究进展综述. 工程科学学报, 2023, 45(10): 1630-1640Wang Ting-Ting, He Xiu-Yu, Zou Yao, Fu Qiang, He Wei. Research progress on the flight control of flapping-wing aerial vehicles. Chinese Journal of Engineering, 2023, 45(10): 1630-1640 [155] 钱辰, 方勇纯, 李友朋. 面向扑翼飞行控制的建模与奇异摄动分析. 自动化学报, 2022, 48(2): 434-443Qian Chen, Fang Yong-Chun, Li You-Peng. Control oriented modeling and singular perturbation analysis in flapping-wing flight. Acta Automatica Sinica, 2022, 48(2): 434-443 -

 下载:
下载:





计量
- 文章访问数: 861
- HTML全文浏览量: 592
- PDF下载量: 291
- 被引次数: 0
