

-
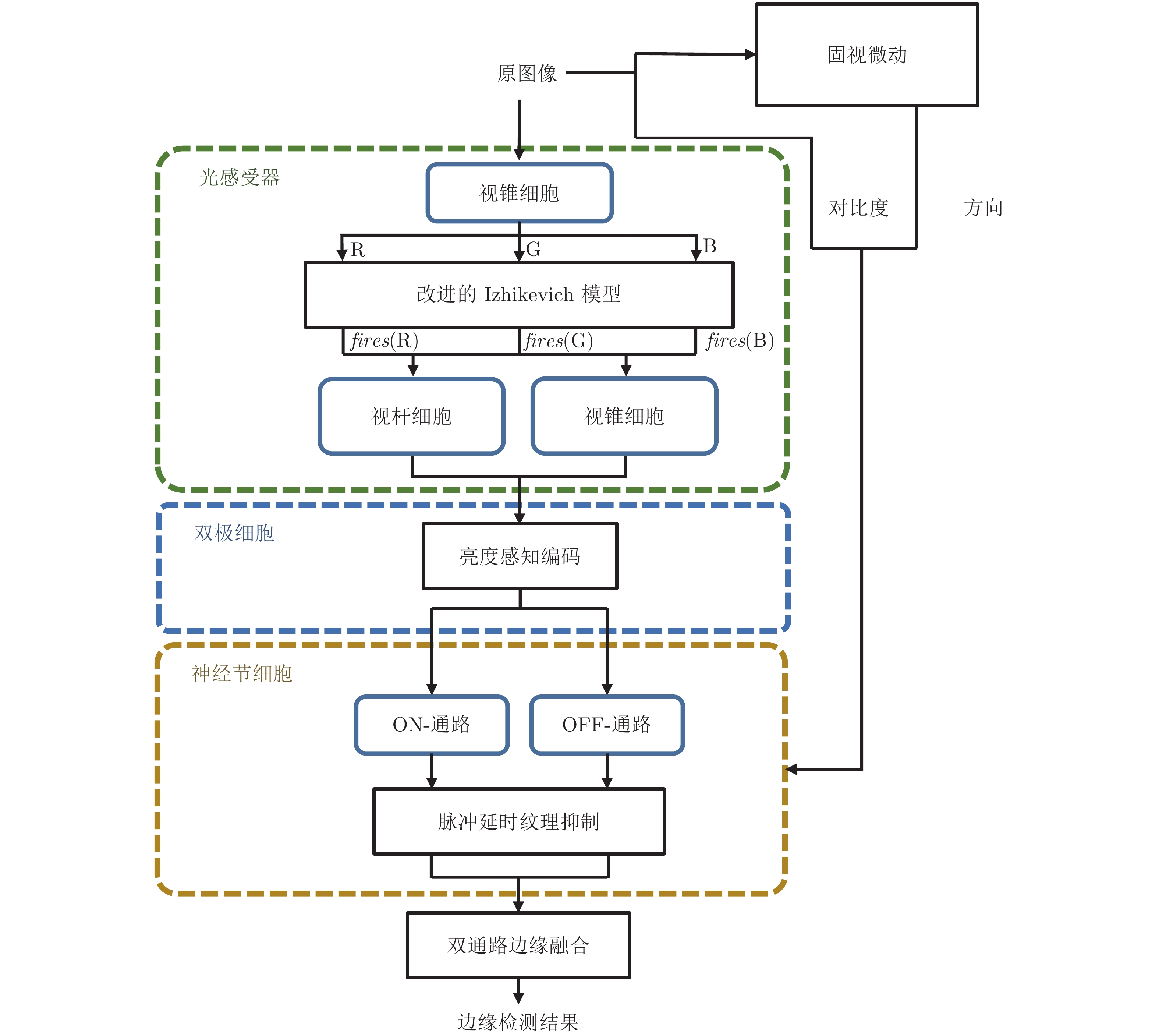
摘要: 基于视网膜对视觉信息的处理方式, 提出一种视网膜功能启发的边缘检测层级模型. 针对视网膜神经元在周期性光刺激下产生适应的特性, 构建具有自适应阈值的Izhikevich神经元模型; 模拟光感受器中视锥细胞、视杆细胞对亮度的感知能力, 构建亮度感知编码层; 引入双极细胞对给光−撤光刺激的分离能力, 并结合神经节细胞对运动方向敏感的特性, 构建双通路边缘提取层; 另外根据神经节细胞神经元在多特征调控下延迟激活的现象, 构建具有脉冲延时特性的纹理抑制层; 最后将双通路边缘提取的结果与延时抑制量相融合, 得到最终边缘检测结果. 以150张来自实验室采集和AGAR数据集中的菌落图像为实验对象对所提方法进行验证, 检测结果的重建图像相似度、边缘置信度、边缘连续性和综合指标分别达到0.9629、0.3111、0.9159和0.7870, 表明所提方法能更有效地进行边缘定位、抑制冗余纹理、保持主体边缘完整性. 本文面向边缘检测任务, 构建了模拟视网膜对视觉信息处理方式的边缘检测模型, 也为后续构建由视觉机制启发的图像计算模型提供了新思路.
-
关键词:
- 边缘检测 /
- 视网膜 /
- Izhikevich模型 /
- 神经编码 /
- 方向选择性神经节细胞
Abstract: Based on the processing of visual information by the retina, this paper proposes a multi-layer model of edge detection inspired by retinal functions. Aiming at the adaptive characteristics of retinal neurons under periodic light stimulation, an Izhikevich neuron model with adaptive threshold is established; By simulating the perception ability of cones and rods for luminance and color in photoreceptors, the luminance perception coding layer is constructed; By introducing the ability of bipolar cells for separating light stimulation, and combining with the characteristics of ganglion cells sensitive to the direction of movement, a multi-pathway edge extraction layer is constructed; In addition, according to the phenomenon of delayed activation of ganglion cell neurons under multi-feature regulation, a texture inhibition layer with pulse delay characteristics is constructed; Finally, by fusing the result of multi-pathway edge extraction with the delay suppression amount, the final edge detection result is obtained. The 150 colony images from laboratory collection and AGAR dataset are used as experimental objects to test the proposed method. The reconstruction image similarity, edge confidence, edge continuity and comprehensive indicators of the detection results are 0.9629, 0.3111, 0.9159 and 0.7870, respectively. The results show that the proposed method can better localize edges, suppress redundant textures, and maintain the integrity of subject edges. This research is oriented to the task of edge detection, constructs an edge detection model that simulates the processing of visual information by the retina, and also provides new ideas for the construction of image computing model inspired by visual mechanism.-
Key words:
- Edge detection /
- retina /
- Izhikevich model /
- neural coding /
- direction-selective ganglion cells (DSGCs)
-

图 2 改进前后的Izhikevich模型对图像进行脉冲发放的结果对比图
Fig. 2 Comparison of the image processing results of the Izhikevich model before and after improvement
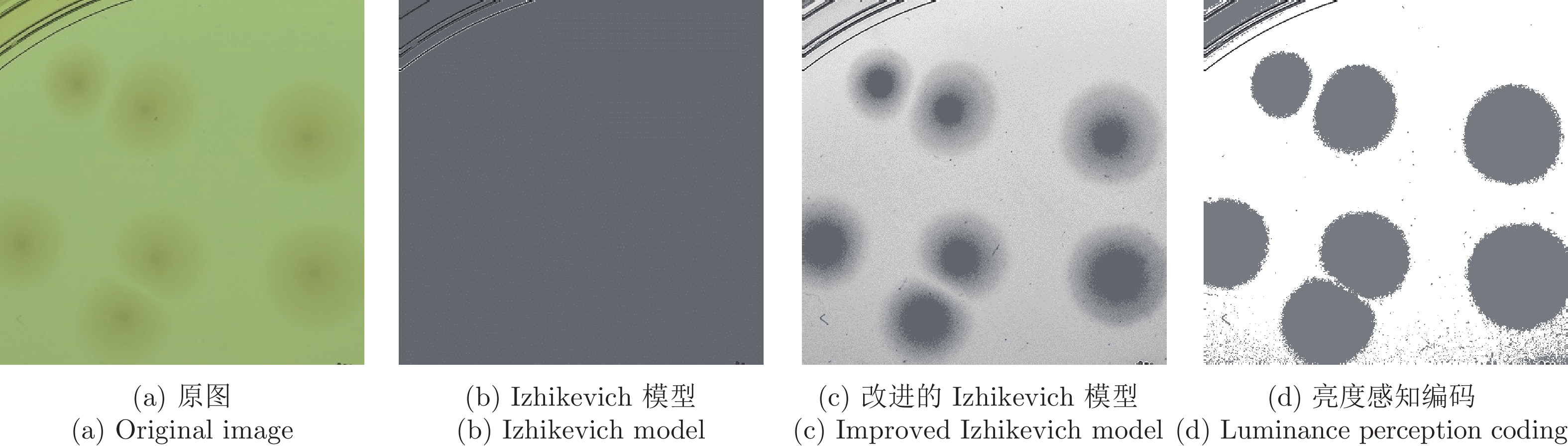
图 3 不同方式对存在弱边缘的菌落图像的处理结果
Fig. 3 Different ways to process the image of colonies with weak edges

图 5 Colony1 ~ Colony5的边缘检测结果(第1行为原图; 第2行为Canny检测的结果; 第3行为SPM检测的结果; 第4行为ISM检测的结果; 第5行为OSM检测的结果; 第6行为NLC检测的结果; 第7行为本文方法检测的结果)
Fig. 5 Edge detection results of Colony1 to Colony5 (The first line is original images; The second line is the results of Canny; The third line is the results of SPM; The fourth line is the results of ISM; The fifth line is the results of OSM; The sixth line is the results of NLC; The seventh line is the results of the proposed method)

图 6 Colony1 ~ Colony5的重建结果 (第1行为原图; 第 2 行为 Canny 检测结果的重建; 第 3 行为 SPM 检测结果的重建; 第 4 行为 ISM 检测结果的重建; 第 5 行为 OSM 检测结果的重建; 第 6 行为 NLC 检测结果的重建; 第 7 行为本文方法检测结果的重建)
Fig. 6 Reconstruction results of Colony1 to Colony5 (The first line is original images; The second line is the results of Canny; The third line is the results of SPM; The fourth line is the results of ISM; The fifth line is the results of OSM; The sixth line is the results of NLC; The seventh line is the results of the proposed method)
表 1 不同检测方法下的重建相似度MSSIM
Table 1 MSSIM of different methods
Serial number MSSIM Canny SPM ISM OSM NLC 本文方法 Colony1 0.7452 0.7725 0.8357 0.9265 0.9175 0.9371 Colony2 0.7951 0.7971 0.8490 0.9528 0.9447 0.9725 Colony3 0.8576 0.8662 0.8314 0.9149 0.8337 0.9278 Colony4 0.9690 0.9827 0.9838 0.9887 0.9893 0.9972 Colony5 0.9634 0.9758 0.9780 0.9771 0.9883 0.9933  下载: 导出CSV
下载: 导出CSV
表 5 不同检测方法下的各评价指标的均值和标准差
Table 5 Average and standard deviation of indexes of different methods
Canny SPM ISM OSM NLC 本文方法 MSSIM 0.8119±0.0649 0.8435±0.0863 0.8692±0.0778 0.9422±0.0291 0.9184±0.0609 0.9629±0.0335 BIdx 0.2091±0.1542 0.1724±0.2056 0.1702±0.1023 0.2763±0.1401 0.2940±0.2221 0.3111±0.1604 CIdx 0.7972±0.0279 0.8101±0.1125 0.8205±0.0561 0.8397±0.2050 0.9149±0.0857 0.9359±0.0306 EIdx 0.6568±0.0562 0.6697±0.0548 0.6801±0.0513 0.7341±0.0281 0.7211±0.0860 0.7870±0.0437
下载: 导出CSV
表 2 不同检测方法下的边缘置信度BIdx
Table 2 BIdx of different methods
Serial number BIdx Canny SPM ISM OSM NLC 本文方法 Colony1 0.4988 0.4618 0.4307 0.5801 0.5058 0.6026 Colony2 0.1821 0.1537 0.1553 0.3365 0.4615 0.4479 Colony3 0.1983 0.1510 0.1610 0.2634 0.1263 0.3257 Colony4 0.1631 0.1488 0.1906 0.1437 0.1521 0.2016 Colony5 0.1620 0.1896 0.1902 0.1882 0.1735 0.1654
下载: 导出CSV
表 3 不同检测方法下的边缘连续性CIdx
Table 3 CIdx of different methods
Serial number CIdx Canny SPM ISM OSM NLC 本文方法 Colony1 0.8377 0.8530 0.8601 0.8676 0.9749 0.9652 Colony2 0.8069 0.8655 0.8533 0.8293 0.9177 0.9518 Colony3 0.8064 0.7408 0.7293 0.8269 0.7764 0.9406 Colony4 0.8143 0.8611 0.9044 0.8430 0.9015 0.9776 Colony5 0.9047 0.8448 0.8632 0.8592 0.8709 0.9571
下载: 导出CSV
表 4 不同检测方法下的综合评价指标EIdx
Table 4 EIdx of different methods
Serial number EIdx Canny SPM ISM OSM NLC 本文方法 Colony1 0.7047 0.7122 0.7344 0.8173 0.8219 0.8473 Colony2 0.7191 0.6508 0.6722 0.7633 0.8244 0.8291 Colony3 0.6048 0.6543 0.6363 0.7258 0.6402 0.7784 Colony4 0.7287 0.7432 0.7649 0.7363 0.7454 0.7690 Colony5 0.7231 0.7454 0.7512 0.7443 0.7262 0.7737
下载: 导出CSV
-
[1] Gong X Y, Su H, Xu D, Zhang Z T, Shen F, Yang H B. An overview of contour detection approaches. International Journal of Automation and Computing, 2018, 15(6): 656-672 doi: 10.1007/s11633-018-1117-z [2] Yedjour H, Meftah B, Lézoray O, Benyettou A. Edge detection based on Hodgkin-Huxley neuron model simulation. Cognitive Processing, 2017, 18(3): 315-323 doi: 10.1007/s10339-017-0803-z [3] 罗佳骏, 武薇, 范影乐, 高云圆. 基于视觉感光层功能的菌落图像多强度边缘检测研究. 中国生物医学工程学报, 2014, 33(6): 677-686Luo Jia-Jun, Wu Wei, Fan Ying-Le, Gao Yun-Yuan. Multi-intensity edge detection for colony images based on the function of photoreceptor in visual system. Chinese Journal of Biomedical Engineering, 2014, 33(6): 677-686 [4] 方芳, 范影乐, 廖进文, 张梦楠. 基于神经元突触可塑性机制图像边缘检测方法. 华中科技大学学报(自然科学版), 2015, (S1): 200−202, 206Fang Fang, Fan Ying-Le, Liao Jin-Wen, Zhang Meng-Nan. Image edge detection method based on synaptic plasticity mechanism. Journal of Huazhong University of Science and Technology (Natural Science Edition), 2015, (S1): 200−202, 206 [5] Fang T, Yuan J T, Yin R, Wu C L. A novel image edge detection method based on the asymmetric STDP mechanism of the visual path. Wireless Communications and Mobile Computing, 2022 [6] Wang B, Chen L L, Zhang Z Y. A novel method on the edge detection of infrared image. Optik, 2019, 180: 610-614 doi: 10.1016/j.ijleo.2018.11.113 [7] Chaoui C N, Ghomari A, Meftah B. Edge and anomaly detection of brain magnetic resonance images in a distributed environment. International Journal of Imaging Systems and Technology, 2022, 32(2): 642-657 doi: 10.1002/ima.22647 [8] Ocko S A, Lindsey J, Ganguli S, Deny S. The emergence of multiple retinal cell types through efficient coding of natural movies. In: Proceedings of the 32nd International Conference on Neural Information Processing Systems. Montreal, Canada: Curran Associates Inc., 2018. 9411−9422 [9] Maheswaranathan N, McIntosh L T, Tanaka H, Grant S, Kastner D B, Melander J B, et al. The dynamic neural code of the retina for natural scenes [Online], available: https://www.biorxiv.org/content/10.1101/340943v5, January 31, 2023 [10] Izhikevich E M. Simple model of spiking neurons. IEEE Transactions on Neural Networks, 2003, 14(6): 1569-1572 doi: 10.1109/TNN.2003.820440 [11] Ruda K, Rudzite A M, Field G D. The functional organization of retinal ganglion cell receptive fields across light levels [Online], available: https://www.biorxiv.org/content/10.1101/2022.09.15.508164v1, January 31, 2023 [12] Ding W, Xiao L, Jing W, Zhang P M, Liang P J. Population activity changes during a trial-to-trial adaptation of bullfrog retinal ganglion cells. Neuroreport, 2014, 25(10): 801-805 doi: 10.1097/WNR.0000000000000191 [13] Hübner C, Schütz A C. Rapid visual adaptation persists across saccades. iScience, 2021, 24(9): Article No. 102986 [14] Li Y C, Cohen E D, Qian H H. Rod and cone coupling modulates photopic ERG responses in the mouse retina. Frontiers in Cellular Neuroscience, 2020, 14: Article No. 566712 [15] Szatko K P, Korympidou M M, Ran Y L, Berens P, Dalkara D, Schubert T, et al. Neural circuits in the mouse retina support color vision in the upper visual field. Nature Communications, 2020, 11: Article No. 3481 [16] Grimes W N, Songco-Aguas A, Rieke F. Parallel processing of rod and cone signals: Retinal function and human perception. Annual Review of Vision Science, 2018, 4: 123-141 doi: 10.1146/annurev-vision-091517-034055 [17] Rucci M, Ahissar E, Burr D. Temporal coding of visual space. Trends in Cognitive Sciences, 2018, 22(10): 883-895 doi: 10.1016/j.tics.2018.07.009 [18] Huang X L, Rangel M, Briggman K L, Wei W. Neural mechanisms of contextual modulation in the retinal direction selective circuit. Nature Communications, 2019, 10: Article No. 2431 [19] Solomon S G. Retinal ganglion cells and the magnocellular, parvocellular, and koniocellular subcortical visual pathways from the eye to the brain. Elsevier, 2021, 178: 31-50 [20] Sugita Y, Miura K, Furukawa T. Retinal ON and OFF pathways contribute to initial optokinetic responses with different temporal characteristics. European Journal of Neuroscience, 2020, 52(4): 3160-3165 doi: 10.1111/ejn.14697 [21] Kerschensteiner D. Feature detection by retinal ganglion cells. Annual Review of Vision Science, 2022, 8: 135-169 doi: 10.1146/annurev-vision-100419-112009 [22] Jacoby J, Schwartz G W. Typology and circuitry of suppressed-by-contrast retinal ganglion cells. Frontiers in Cellular Neuroscience, 2018, 12: Article No. 00269 [23] Iacaruso M F, Gasler I T, Hofer S B. Synaptic organization of visual space in primary visual cortex. Nature, 2017, 547(7664): 449-452 doi: 10.1038/nature23019 [24] 廖进文, 范影乐, 武薇, 高云圆, 李轶. 基于抑制性突触多层神经元群放电编码的图像边缘检测. 中国生物医学工程学报, 2014, 33(5): 513-524 doi: 10.3969/j.issn.0258-8021.2014.05.01Liao Jin-Wen, Fan Ying-Le, Wu Wei, Gao Yun-yuan, Li Yi. Image edge detection based on spike coding of multilayer neuronal population with inhibitory synapse. Chinese Journal of Biomedical Engineering, 2014, 33(5): 513-524 doi: 10.3969/j.issn.0258-8021.2014.05.01 [25] 王典, 范影乐, 张梦楠, 武薇. 基于突触连接视通路方向敏感的图像分级边缘检测方法. 中国生物医学工程学报, 2015, 34(5): 522-532Wang Dian, Fan Ying-Le, Zhang Meng-Nan, Wu Wei. A hierarchical image edge detection method based on orientation sensitivity of visual pathway with synaptic connections. Chinese Journal of Biomedical Engineering, 2015, 34(5): 522-532 [26] Majchrowska S, Pawlowski J, Gula G, Bonus T, Hanas A, Loch A, et al. AGAR a microbial colony dataset for deep learning detection. arXiv preprint arXiv: 2108.01234, 2021. [27] Govindarajan B, Panetta K A, Agaian S. Image reconstruction for quality assessment of edge detectors. In: Proceedings of the IEEE International Conference on Systems, Man and Cybernetics. Singapore: IEEE, 2008. 691−696 [28] Meer P, Georgescu B. Edge detection with embedded confidence. IEEE Transactions on Pattern Analysis and Machine Intelligence, 2001, 23(12): 1351-1365 doi: 10.1109/34.977560 [29] 磨少清. 边缘检测及其评价方法的研究 [博士学位论文], 天津大学, 中国, 2011Mo Shao-Qing. Research on Edge Detection and Its Evaluation [Ph.D. dissertation], Tianjin University, China, 2011 [30] Niell C M, Scanziani M. How cortical circuits implement cortical computations: Mouse visual cortex as a model. Annual Review of Neuroscience, 2021, 44: 517-546 doi: 10.1146/annurev-neuro-102320-085825 [31] Huang X L, Kim A J, Ledesma H A, Ding J, Smith R G, Wei W. Visual stimulation induces distinct forms of sensitization of On-Off direction-selective ganglion cell responses in the dorsal and ventral retina. Journal of Neuroscience, 2022, 42(22): 4449-4469 doi: 10.1523/JNEUROSCI.1391-21.2022 [32] Kupers E R, Benson N C, Carrasco M, Winawer J. Asymmetries around the visual field: From retina to cortex to behavior. PLoS Computational Biology, 2022, 18(1): Article No. e1009771 -

 下载:
下载:



计量
- 文章访问数: 847
- HTML全文浏览量: 599
- PDF下载量: 194
- 被引次数: 0
