

-
摘要:
集群是鱼类生物中一种常见的现象, 特定编队的集群运动可以显著提高鱼群的游动效率. 鱼集群游动节能机理的研究为机器人集群编队设计和控制提供启发与帮助, 得到了研究人员的广泛关注. 本文介绍了鱼集群游动节能机理研究的主要方法及最新的研究成果, 将研究方法分为鱼群观察分析法、计算流体力学仿真法和实验装置研究法, 并基于此对近些年的研究成果进行了综述和分析, 最后列举了鱼集群游动节能机理研究的主要问题与未来发展方向.
Abstract:Schooling is a common phenomenon in fish creatures. The swimming efficiency of fish can be significantly improved by schooling movement of specific formation. The research on the energy-saving mechanism of fish schooling provides inspiration and help for the design and control of robot swarm formation, and has been widely concerned by researchers. In this paper, the main methods and the latest research results of energy-saving mechanism of fish schooling are introduced. We divide the research methods into three categories, namely observation analysis approach, computational fluid dynamics simulation approach, and experimental setup research approach. Achievements of the research on the energy-saving mechanism based on these methods are discussed thoroughly. Finally, the main problem and future research directions are listed.
-
Key words:
- Fish schooling /
- cluster swimming /
- energy-saving mechanism /
- formation
-
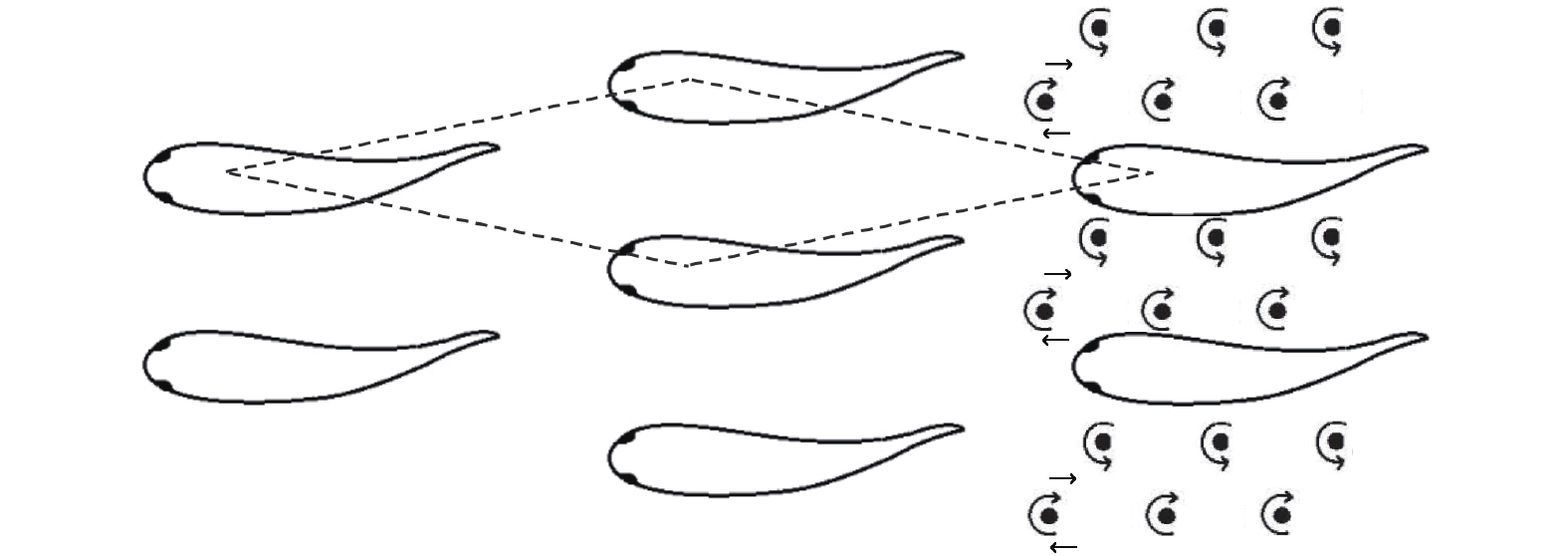

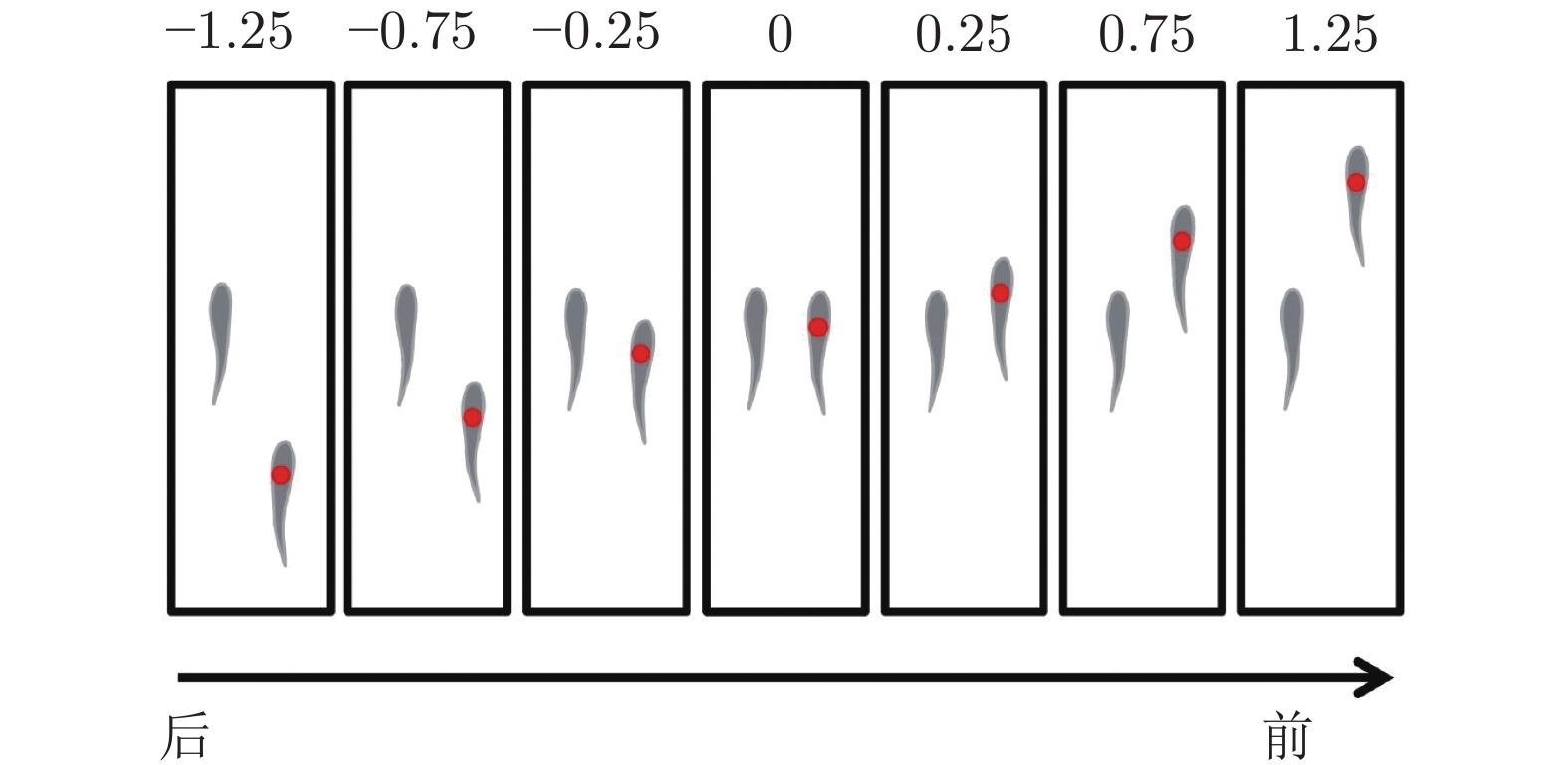
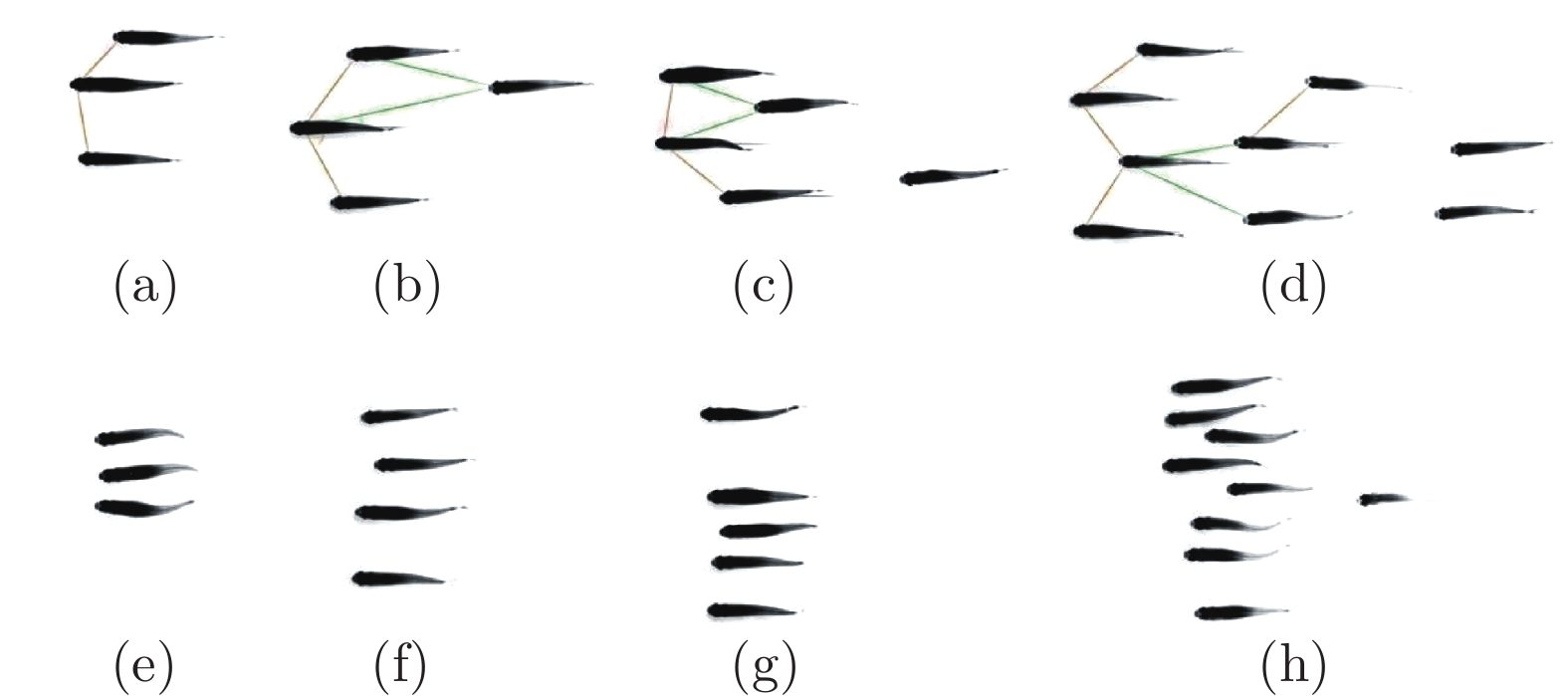


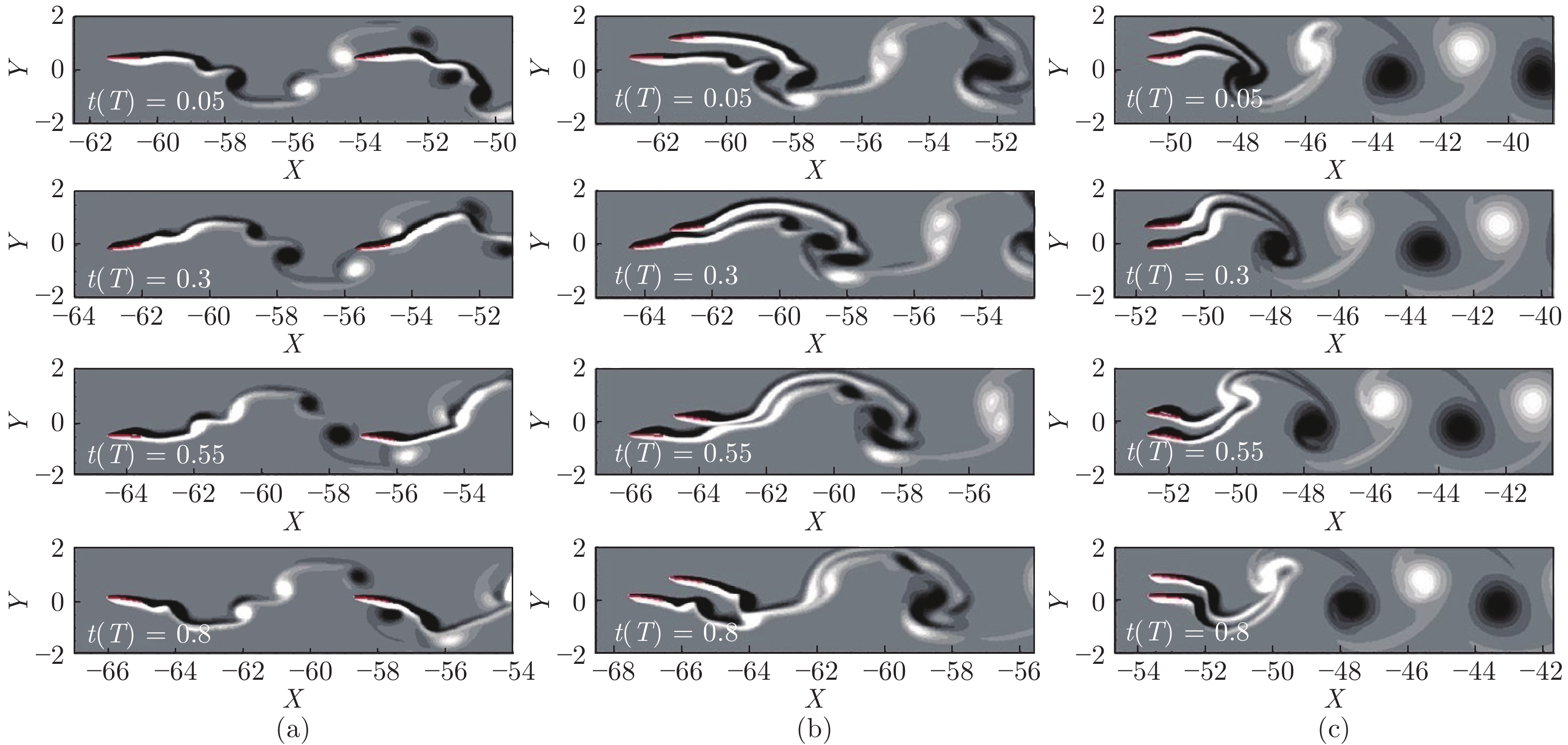

图 11 由两条、三条和四条鱼组成的队列及涡度结构示意图 ((a)两鱼并排(反相位); (b)两鱼并排(同相); (c)两鱼串列(紧凑); (d)两鱼串列(松散); (e)两鱼交错(紧凑); (f)两鱼交错(松散); (g)三鱼并排(反相位); (h)三鱼并排(同相); (i)三鱼梯队; (j)三鱼交错(I型); (k)三条鱼交错(II型); (l)四条鱼矩形(紧凑, 反相位); (m)四条鱼矩形(松散, 反相位); (n)四条鱼菱形)[72]
Fig. 11 The swarm configurations and flow structures of two, three and four fish ((a) two fish side-by-side (anti-phase); (b) two fish side-by-side (in-phase); (c) two fish in-line (compact); (d) two fish in-line (loose); (e) two fish staggered (compact); (f) two fish staggered (loose); (g) three fish side-by-side (anti-phase); (h) three fish side-by-side (in-phase); (i) three fish echelon; (j) three fish staggered (type I); (k) three fish staggered (type II); (l) four fish rectangular (compact, anti-phase); (m) four fish rectangular (loose, anti-phase); (n) four fish diamond)[72]
表 1 鱼群观察分析法的发展历程
Table 1 The development of the method of observation and analysis for fish schooling
作者 研究方法 主要结论 (涡流效应) Brede[18] (1965) 观察鱼群游动过程 维持漩涡的完整性对鱼群游动的效率很重要 Weihs[1, 19] (1973) 观察鱼群, 构建二维鱼群编队水动力模型 菱形为最优队形, 鱼群有效利用漩涡最多可节省50%的能量 Herskin等[10] (1998) 测量9条海鲈鱼集群游动时的尾拍频率 后方鲈鱼的尾拍频率比在前方鲈鱼最多降低14%, 耗氧率降低23% Svendsen等[21] (2003) 测量8条拟鲤组成菱形编队游动时的尾拍频率 后方鱼的尾拍频率最多可比在前方低11.9%, 菱形编队有利于节能 Liao等[22-25] (2003) 观察在圆柱挡板后游动的鳟鱼, DPIV观察涡流分布, 生物肌电信号测量仪测量鳟鱼的肌电信号 鳟鱼通过感知并利用涡流来保持卡门步态, 减少了肌肉活动. 证明鳟鱼会自动改变身体运动来从漩涡中获取能量 作者 研究方法 主要结论 (槽道效应) Burgerhout等[31] (2013) 观察7条欧洲鳗鲡在水槽中集群游动过程, 并测量鳗鱼尾拍频率 鳗鱼集群游动可减少30%功耗. 鳗鱼倾向于以彼此平行的同步方式游动, 利用个体之间的侧向力达到节能目的 Marras等[35] (2015) 利用相机与DPIV观察灰鲻鱼集群游动及水流情况, 测量标记鱼在不同位置的尾拍频率与幅值 鱼群任何位置都可以节能, 耗氧率最多降低19.4%. 不只是尾部涡流, 鱼体前端周围的流体动力学也有助于提高邻近鱼游动效率 Ashraf等[36-37] (2016, 2017) 利用立体视频记录仪跟踪红鼻四头鱼集群中每条鱼的3D位置及尾鳍摆动 鱼群快速游动时, 倾向于“矩形”或“并列”编队. 鱼间的水动力相互作用可以提高游动效率, 证明槽道效应在鱼群节能过程中发挥重要作用  下载: 导出CSV
下载: 导出CSV
表 2 CFD仿真法的发展历程
Table 2 The development of the method of CFD simulation for fish schooling
作者 研究方法 主要结论 (涡流效应) Kelly等[44-45] (2005, 2010),
Deng等[46] (2006), Chung[47] (2011)从菱形鱼群中提取出三条鱼作为基本单元, 对其游动进行数值模拟 后方鱼可捕获前方鱼制造的尾涡, 并从脱落的反卡门涡街中受益达到节能目的 Pan等[48] (2010), Shao等[49] (2010),
Xiao等[50-51] (2011, 2012),
Chao[52-53] (2018, 2019)数值模拟D形圆柱后柔性板NACA0012的被动运动 上游圆柱的存在增强了其尾流中的反卡门涡街, 后方柔性板的推力系数可提高4倍 Khalid等[54] (2016), Tian等[55-56]
(2011, 2016),
Maertens等[57] (2017)分别数值模拟两条二维与三维的仿真鱼在编队中的游动 串联和交错编队均能提高游动效率, 后方鱼效率最高可提高30%. 通过利用迎面漩涡, 后方鱼可在能与前方鱼尾迹相互作用的位置受益 Novati等[62] (2017), Verma等[63] (2018) 分别数值模拟两条二维与三维自主仿真鱼编队游动, 先导鱼步态固定, 尾随鱼利用深度强化学习来调整步态 尾随鱼的能量消耗减少30%, 游动效率增加20%. 鱼类可以通过将身体置于前方鱼身后的适当位置并拦截其脱落的涡流来提高自身推进效率 作者 研究方法 主要结论 (槽道效应) 王亮[64] (2007) 数值模拟多条二维仿真鱼在粘性流体中编队游动 小鱼跟随大鱼多利用侧向漩涡来提高推进效率. 体形相差不大的鱼多利用尾涡来提高的推进效率 Daghooghi等[14] (2015) 数值模拟多条三维仿真鱼以矩形编队自主同步游动 同等能量消耗下集群游动的速度比单独游动快20%. 鱼间距离减小会提高游动方向的流体速度, 单靠流体动力相互作用足以产生高效的集群游动 Hemelrijk等[66] (2015),
Li 等[67] (2019)利用数值模拟研究了鱼群四种不同编队(串列、方阵、菱形和矩形)游动时的Froude效率 鱼集群游动比单独游动效率高, 但菱形队列并不是最优. 尾流和横向压力共同影响游动效率, 尾流主要影响推力, 横向压力主要影响功率损失 Dai等[72] (2018) 分别数值模拟了2、3和4条仿真鱼组成的编队自主游动, 并用运输成本来量化稳定编队的游动效率 与单独游动相比, 集群游动COT最多减少16%, 但相较于其他编队, 菱形编队并没有表现出任何节能优势, 说明被动水动力相互作用是节能的主要原因
下载: 导出CSV
表 3 实验装置研究法的发展历程
Table 3 The development of the method of experimental setup research for fish schooling
作者 研究方法 主要结论 Dewey等[79-80]
(2013, 2014),
Boschitsch等[81] (2014)利用高速相机及DPIV观察分析两个仿生柔性水翼在并列与串列结构中的摆动 并联结构中同相摆动尾流形成涡对, 诱导动量喷流来提高水翼推进效率; 串联结
构中尾流形成相干模式, 后方水翼的效率最多可提高50%Becker等[83] (2015) 扑翼阵自主推进实验 仅通过流体动力学相互作用足以产生互相耦合的集群运动, 来提高推进效率 裴正楷等[84-85] (2016) 研究两条仿鲹科机器鱼并排运动 通过流体传导, 并排游动的机器鱼在特定相位差时可以相互促进, 提高运动效率
下载: 导出CSV
表 4 三类研究方法特点对比
Table 4 Comparison of three kinds of research methods
对比项目 优点 缺点 鱼群观察分析法 反映真实鱼群游动情况 编队不易保持, 不易精确定量分析 计算流体力学仿真法 可精确定量分析, 模拟各种编队 不能反映真实的水动力学关系 实验装置研究法 可反映真实水动力学关系, 易保持队形, 可定量分析 无法完全模拟鱼游动过程, 对实验装置和平台要求较高
下载: 导出CSV
-
[1] Weihs D. Hydromechanics of fish schooling. Nature, 1973, 241(5387): 290−291 doi: 10.1038/241290a0 [2] Shaw E. Schooling fishes. American Scientist, 1978, 66(2): 166−175 [3] Pitcher T J, Parrish J K. Functions of shoaling behavior in teleosts. Behavior of Teleost Fishes (2nd edition). London: Chapman and Hall, 1993: 363−439 [4] Godin J G J, Morgan M J. Predator avoidance and school size in a cyprinodontid fish, the banded killifish (Fundulus diaphanous lesueur). Behavioral Ecology and Sociobiology, 1985, 16(2): 105−110 doi: 10.1007/BF00295142 [5] Magurran A E, Higham A. Information transfer across fish shoals under predator threat. Ethology, 1988, 78(2): 153−158 [6] Pitcher T J, Magurran A E, Winfield I J. Fish in larger shoals find food faster. Behavioral Ecology and Sociobiology, 1982, 10(2): 149−151 doi: 10.1007/BF00300175 [7] Wolf N G. Schooling tendency and foraging benefit in the ocean surgeonfish. Behavioral Ecology and Sociobiology, 1987, 21(1): 59−63 doi: 10.1007/BF00324436 [8] Ranta E, Lindström K. Assortative schooling in three-spined sticklebacks? Annales Zoologici Fennici, 1990, 27(2): 67−75 [9] Fish F. Energetics of swimming and flying in formation. Theoretical Biology, 1999, 5: 283−304 [10] Herskin J, Steffensen J F. Energy savings in sea bass swimming in a school: Measurements of tail beat frequency and oxygen consumption at different swimming speeds. Journal of Fish Biology, 1998, 53(2): 366−376 doi: 10.1111/j.1095-8649.1998.tb00986.x [11] Johansen J L, Vaknin R, Steffensen J F, Domenici P. Kinematics and energetic benefits of schooling in the labriform fish, striped surfperch Embiotoca lateralis. Marine Ecology Progress Series, 2010, 420: 221−229 doi: 10.3354/meps08885 [12] Killen S S, Marras S, Steffensen J F, McKenzie D J. Aerobic capacity influences the spatial position of individuals within fish schools. Proceedings of the Royal Society B: Biological Sciences, 2012, 279(1727): 357−364 doi: 10.1098/rspb.2011.1006 [13] Ross R M, Backman T W H, Limburg K E. Notes: Group-size-mediated metabolic rate reduction in American shad. Transactions of the American Fisheries Society, 1992, 121(3): 385−390 doi: 10.1577/1548-8659(1992)121<0385:NGMRRI>2.3.CO;2 [14] Daghooghi M, Borazjani I. The hydrodynamic advantages of synchronized swimming in a rectangular pattern. Bioinspiration & Biomimetics, 2015, 10(5): 056018 [15] Delcourt J, Denoël M, Ylieff M, Poncin P. Video multitracking of fish behaviour: A synthesis and future perspectives. Fish and Fisheries, 2013, 14(2): 186−204 doi: 10.1111/j.1467-2979.2012.00462.x [16] Lighthill M J. Large-amplitude elongated-body theory of fish locomotion. Proceedings of the Royal Society B: Biological Sciences, 1971, 179(1055): 125−138 [17] Wen L, Wang T M, Wu G H, Liang J H. Quantitative thrust efficiency of a self-propulsive robotic fish: Experimental method and hydrodynamic investigation. IEEE/ASME Transactions on Mechatronics, 2013, 18(3): 1027−1038 doi: 10.1109/TMECH.2012.2194719 [18] Breder C M. Vortices and fish schools. Zoologica: Scientific Contributions of the New York Zoological Society, 1965, 50(10): 97−114 [19] Weihs D. Optimal fish cruising speed. Nature, 1973, 245(5419): 48−50 doi: 10.1038/245048a0 [20] Fish F E, Fegely J F, Xanthopoulos C J. Burst-and-coast swimming in schooling fish (Notemigonus crysoleucas) with implications for energy economy. Comparative Biochemistry and Physiology Part A: Physiology, 1991, 100(3): 633−637 doi: 10.1016/0300-9629(91)90382-M [21] Svendsen J C, Skov J, Bildsoe M, Steffensen J F. Intra-school positional preference and reduced tail beat frequency in trailing positions in schooling roach under experimental conditions. Journal of Fish Biology, 2003, 62(4): 834−846 doi: 10.1046/j.1095-8649.2003.00068.x [22] Liao J C, Beal D N, Lauder G V, Triantafyllou M S. Fish exploiting vortices decrease muscle activity. Science, 2003, 302(5650): 1566−1569 doi: 10.1126/science.1088295 [23] Liao J C, Beal D N, Lauder G V, Triantafyllou M S. The Kármán gait: Novel body kinematics of rainbow trout swimming in a vortex street. Journal of Experimental Biology, 2003, 206(6): 1059−1073 doi: 10.1242/jeb.00209 [24] Liao J C. Neuromuscular control of trout swimming in a vortex street: Implications for energy economy during the Karman gait. Journal of Experimental Biology, 2004, 207(20): 3495−3506 doi: 10.1242/jeb.01125 [25] Liao J C. The role of the lateral line and vision on body kinematics and hydrodynamic preference of rainbow trout in turbulent flow. Journal of Experimental Biology, 2006, 209(20): 4077−4090 doi: 10.1242/jeb.02487 [26] Marras S, Porfiri M. Fish and robots swimming together: Attraction towards the robot demands biomimetic locomotion. Journal of the Royal Society Interface, 2012, 9(73): 1856−1868 doi: 10.1098/rsif.2012.0084 [27] Halsey L G, Wright S, Racz A, Metcalfe J D, Killen S S. How does school size affect tail beat frequency in turbulent water? Comparative Biochemistry and Physiology Part A: Molecular & Integrative Physiology, 2018, 218: 63−69 [28] Lopez U, Gautrais J, Couzin I D, Theraulaz G. From behavioural analyses to models of collective motion in fish schools. Interface Focus, 2012, 2(6): 693−707 doi: 10.1098/rsfs.2012.0033 [29] Liao J C. A review of fish swimming mechanics and behaviour in altered flows. Philosophical Transactions of the Royal Society B: Biological Sciences, 2007, 362(1487): 1973−1993 doi: 10.1098/rstb.2007.2082 [30] Webb P W. The effect of solid and porous channel walls on steady swimming of steelhead trout oncorhynchus mykiss. Journal of Experimental Biology, 1993, 178(1): 97−108 [31] Burgerhout E, Tudorache C, Brittijn S A, Palstra A P, Dirks R P, van den Thillart G E E J M. Schooling reduces energy consumption in swimming male European eels, Anguilla anguilla L. Journal of Experimental Marine Biology and Ecology, 2013, 448: 66−71 doi: 10.1016/j.jembe.2013.05.015 [32] Kern S, Koumoutsakos P. Simulations of optimized anguilliform swimming. Journal of Experimental Biology, 2006, 209(24): 4841−4857 doi: 10.1242/jeb.02526 [33] Tytell E D, Lauder G V. The hydrodynamics of eel swimming. I. Wake structure. Journal of Experimental Biology, 2004, 207(11): 1825−1841 doi: 10.1242/jeb.00968 [34] Lauder G V, Tytell E D. Hydrodynamics of undulatory propulsion. Fish Physiology, 2005, 23: 425−468 [35] Marras S, Killen S S, Lindström J, McKenzie D J, Steffensen J F, Domenici P. Fish swimming in schools save energy regardless of their spatial position. Behavioral Ecology and Sociobiology, 2015, 69(2): 219−226 doi: 10.1007/s00265-014-1834-4 [36] Ashraf I, Godoy-Diana R, Halloy J, Collignon B, Thiria B. Synchronization and collective swimming patterns in fish (Hemigrammus bleheri). Journal of the Royal Society Interface, 2016, 13(123): 20160734 doi: 10.1098/rsif.2016.0734 [37] Ashraf I, Bradshaw H, Ha T T, Halloy J, Godoy-Diana R, Thiria B. Simple phalanx pattern leads to energy saving in cohesive fish schooling. Proceedings of the National Academy of Sciences of the United States of America, 2017, 114(36): 9599−9604 doi: 10.1073/pnas.1706503114 [38] Peskin C S. Flow patterns around heart valves: A numerical method. Journal of Computational Physics, 1972, 10(2): 252−271 doi: 10.1016/0021-9991(72)90065-4 [39] Triantafyllou M S, Triantafyllou G S. An efficient swimming machine. Scientific American, 1995, 272(3): 64−70 doi: 10.1038/scientificamerican0395-64 [40] Maertens A P, Triantafyllou M S, Yue D K P. Efficiency of fish propulsion. Bioinspiration & Biomimetics, 2015, 10(4): 046013 [41] Borazjani I, Sotiropoulos F. Numerical investigation of the hydrodynamics of carangiform swimming in the transitional and inertial flow regimes. Journal of Experimental Biology, 2008, 211(10): 1541−1558 doi: 10.1242/jeb.015644 [42] Borazjani I, Sotiropoulos F. Numerical investigation of the hydrodynamics of anguilliform swimming in the transitional and inertial flow regimes. Journal of Experimental Biology, 2009, 212(4): 576−592 doi: 10.1242/jeb.025007 [43] Borazjani I, Sotiropoulos F. On the role of form and kinematics on the hydrodynamics of self-propelled body/caudal fin swimming. Journal of Experimental Biology, 2010, 213(1): 89−107 doi: 10.1242/jeb.030932 [44] Kelly S D, Xiong H L, Controlled hydrodynamic interactions in schooling aquatic locomotion. In: Proceedings of the 44th IEEE Conference on Decision and Control. Seville, Spain: IEEE, 2005. 3904−3910 [45] Kelly S D, Pujari P. Propulsive energy harvesting by a fishlike vehicle in a vortex flow: Computational modeling and control. In: Proceedings of the 49th IEEE Conference on Decision and Control. Atlanta, USA: IEEE, 2010. 1058−1064 [46] Deng J, Shao X M. Hydrodynamics in a diamond-shaped fish school. Journal of Hydrodynamics, Ser. B, 2006, 18(3S): 438−442 [47] Chung M H. Hydrodynamic performance of two-dimensional undulating foils in triangular formation. Journal of Mechanics, 2011, 27(2): 177−190 doi: 10.1017/jmech.2011.21 [48] Pan D Y, Liu H, Shao X M. Studies on the oscillation behavior of a flexible plate in the wake of a D-cylinder. Journal of Hydrodynamics, 2010, 22(S1): 132−137 doi: 10.1016/S1001-6058(09)60182-7 [49] Shao X M, Pan D Y, Deng J, Yu Z S. Hydrodynamic performance of a fishlike undulating foil in the wake of a cylinder. Physics of Fluids, 2010, 22(11): 111903 doi: 10.1063/1.3504651 [50] Xiao Q, Sun K, Liu H, Hu J X. Computational study on near wake interaction between undulation body and a D-section cylinder. Ocean Engineering, 2011, 38(4): 673−683 doi: 10.1016/j.oceaneng.2010.12.017 [51] Xiao Q, Liu W D, Hu J X. Parametric study on a cylinder drag reduction using downstream undulating foil. European Journal of Mechanics — B/Fluids, 2012, 36: 48−62 doi: 10.1016/j.euromechflu.2012.04.005 [52] Chao L M, Zhang D, Cao Y H, Pan G. Numerical studies on the interaction between two parallel D-cylinder and oscillated foil. Modern Physics Letters B, 2018, 32(6): 1850034 doi: 10.1142/S0217984918500343 [53] Chao L M, Pan G, Zhang D, Yan G X. On the thrust generation and wake structures of two travelling-wavy foils. Ocean Engineering, 2019, 183: 167−174 doi: 10.1016/j.oceaneng.2019.04.097 [54] Khalid M S U, Akhtar I, Dong H B. Hydrodynamics of a tandem fish school with asynchronous undulation of individuals. Journal of Fluids and Structures, 2016, 66: 19−35 doi: 10.1016/j.jfluidstructs.2016.07.008 [55] Tian F B, Wang W Q, Wu J, Sui Y. Swimming performance and vorticity structures of a mother-calf pair of fish. Computers & Fluids, 2016, 124: 1−11 [56] Tian F B, Luo H X, Zhu L D, Liao J C, Lu X Y. An efficient immersed boundary-lattice Boltzmann method for the hydrodynamic interaction of elastic filaments. Journal of Computational Physics, 2011, 230(19): 7266−7283 doi: 10.1016/j.jcp.2011.05.028 [57] Maertens A P, Gao A, Triantafyllou M S. Optimal undulatory swimming for a single fish-like body and for a pair of interacting swimmers. Journal of Fluid Mechanics, 2017, 813: 301−345 doi: 10.1017/jfm.2016.845 [58] Li G, Kolomenskiy D, Liu H, Thiria B, Godoy-Diana R. On the energetics and stability of a minimal fish school. PLoS One, 2019, 14(8): e0215265 doi: 10.1371/journal.pone.0215265 [59] Gazzola M, Tchieu A A, Alexeev D, de Brauer A, Koumoutsakos P. Learning to school in the presence of hydrodynamic interactions. Journal of Fluid Mechanics, 2016, 789: 726−749 doi: 10.1017/jfm.2015.686 [60] Wolfgang M J, Anderson J M, Grosenbaugh M A, Yue D K P, Triantafyllou M S. Near-body flow dynamics in swimming fish. The Journal of Experimental Biology, 1999, 202(17): 2303−2327 [61] Tchieu A A, Kanso E, Newton P K. The finite-dipole dynamical system. Proceedings of the Royal Society A, 2012, 468(2146): 3006−3026 doi: 10.1098/rspa.2012.0119 [62] Novati G, Verma S, Alexeev D, Rossinelli D, van Rees W M, Koumoutsakos P. Synchronisation through learning for two self-propelled swimmers. Bioinspiration & Biomimetics, 2017, 12(3): 036001 [63] Verma S, Novati G, Koumoutsakos P. Efficient collective swimming by harnessing vortices through deep reinforcement learning. Proceedings of the National Academy of Sciences of the United States of America, 2018, 115(23): 5849−5854 doi: 10.1073/pnas.1800923115 [64] 王亮. 仿生鱼群自主游动及控制的研究[博士学位论文], 河海大学, 中国, 2007.Wang Liang. Numerical Simulation and Control of Self-propelled Swimming of Bionics Fish School [Ph.D. dissertation], Hohai University, China, 2007. [65] Dong G J, Lu X Y. Characteristics of flow over traveling wavy foils in a side-by-side arrangement. Physics of Fluids, 2007, 19(5): 057107 doi: 10.1063/1.2736083 [66] Hemelrijk C K, Reid D A P, Hildenbrandt H, Padding J T. The increased efficiency of fish swimming in a school. Fish and Fisheries, 2015, 16(3): 511−521 doi: 10.1111/faf.12072 [67] Li S M, Li C, Xu L Y, Yang W J, Chen X C. Numerical simulation and analysis of fish-like robots swarm. Applied Sciences, 2019, 9(8): 1652 doi: 10.3390/app9081652 [68] 谢春梅, 黄伟希. 前后排列柔性细丝在黏性流体中自主推进的稳定形态及动力学机制. 科学通报, 2017, 62(19): 2094−2103 doi: 10.1360/N972016-01312Xie Chun-Mei, Huang Wei-Xi. Stable states and mechanism of self-propulsion of two tandem filaments in viscous flow. Chinese Science Bulletin, 2017, 62(19): 2094−2103 doi: 10.1360/N972016-01312 [69] Peng Z R, Huang H B, Lu X Y. Collective locomotion of two closely spaced self-propelled flapping plates. Journal of Fluid Mechanics, 2018, 849: 1068−1095 doi: 10.1017/jfm.2018.447 [70] Peng Z R, Huang H B, Lu X Y. Hydrodynamic schooling of multiple self-propelled flapping plates. Journal of Fluid Mechanics, 2018, 853: 587−600 doi: 10.1017/jfm.2018.634 [71] Peng Z R, Huang H B, Lu X Y. Collective locomotion of two self-propelled flapping plates with different propulsive capacities. Physics of Fluids, 2018, 30(11): 111901 doi: 10.1063/1.5053662 [72] Dai L Z, He G W, Zhang X, Zhang X. Stable formations of self-propelled fish-like swimmers induced by hydrodynamic interactions. Journal of the Royal Society Interface, 2018, 15(147): 20180490 doi: 10.1098/rsif.2018.0490 [73] Lin X J, Wu J, Zhang T W, Yang L M. Self-organization of multiple self-propelling flapping foils: Energy saving and increased speed. Journal of Fluid Mechanics, 2020, 884: R1 doi: 10.1017/jfm.2019.954 [74] Lin X J, He G Y, He X Y, Wang Q, Chen L S. Hydrodynamic studies on two wiggling hydrofoils in an oblique arrangement. Acta Mechanica Sinica, 2018, 34(3): 446−451 doi: 10.1007/s10409-017-0732-1 [75] Lin X J, He G Y, He X Y, Wang Q. Dynamic response of a semi-free flexible filament in the wake of a flapping foil. Journal of Fluids and Structures, 2018, 83: 40−53 doi: 10.1016/j.jfluidstructs.2018.08.009 [76] Lin X J, Wu J, Zhang T W, Yang L M. Phase difference effect on collective locomotion of two tandem autopropelled flapping foils. Physical Review Fluids, 2019, 4(5): 054101 doi: 10.1103/PhysRevFluids.4.054101 [77] Park S G, Sung H J. Hydrodynamics of flexible fins propelled in tandem, diagonal, triangular and diamond configurations. Journal of Fluid Mechanics, 2018, 840: 154−189 doi: 10.1017/jfm.2018.64 [78] Chen S Y, Fei Y H J, Chen Y C, Chi K J, Yang J T. The swimming patterns and energy-saving mechanism revealed from three fish in a school. Ocean Engineering, 2016, 122: 22−31 doi: 10.1016/j.oceaneng.2016.06.018 [79] Dewey P A, Boschitsch B M, Moored K W, Stone H A, Smits A J. Scaling laws for the thrust production of flexible pitching panels. Journal of Fluid Mechanics, 2013, 732: 29−46 doi: 10.1017/jfm.2013.384 [80] Dewey P A, Quinn D B, Boschitsch B M, Smits A J. Propulsive performance of unsteady tandem hydrofoils in a side-by-side configuration. Physics of Fluids, 2014, 26(4): 041903 doi: 10.1063/1.4871024 [81] Boschitsch B M, Dewey P A, Smits A J. Propulsive performance of unsteady tandem hydrofoils in an in-line configuration. Physics of Fluids, 2014, 26(5): 051901 doi: 10.1063/1.4872308 [82] Ryuh Y S, Yang G H, Liu J D, Hu H S. A school of robotic fish for mariculture monitoring in the sea coast. Journal of Bionic Engineering, 2015, 12(1): 37−46 doi: 10.1016/S1672-6529(14)60098-6 [83] Becker A D, Masoud H, Newbolt J W, Shelley M, Ristroph L. Hydrodynamic schooling of flapping swimmers. Nature Communications, 2015, 6: 8514 doi: 10.1038/ncomms9514 [84] 裴正楷, 李亮, 陈世明, 潘想, 谢广明. 机器鱼在线功率检测系统设计与实现. 测控技术, 2016, 35(11): 9−13 doi: 10.3969/j.issn.1000-8829.2016.11.003Pei Zheng-Kai, Li Liang, Chen Shi-Ming, Pan Xiang, Xie Guang-Ming. Design and implementation of online power capture system for robotic fish. Measurement and Control Technology, 2016, 35(11): 9−13 doi: 10.3969/j.issn.1000-8829.2016.11.003 [85] 裴正楷, 刘俊恺, 陈世明, 李亮. 双鱼并排游动时水动力性能研究. 测控技术, 2016, 35(12): 16−20 doi: 10.3969/j.issn.1000-8829.2016.12.004Pei Zheng-Kai, Liu Jun-Kai, Chen Shi-Ming, Li Liang. Hydrodynamic performance of Beas swimming side by side. Measurement & Control Technology, 2016, 35(12): 16−20 doi: 10.3969/j.issn.1000-8829.2016.12.004 [86] Zhang Z, Yang T, Zhang T H, Zhou F H, Cen N, Li T F, et al. Global vision-based formation control of soft robotic fish swarm. Soft Robotics, 2020. DOI: 10.1089/soro.2019.0174 -

 下载:
下载:



计量
- 文章访问数: 1540
- HTML全文浏览量: 1745
- PDF下载量: 545
- 被引次数: 0
