

Time Correlation of Time-invariant Linear Models in Neural Decoding for the Macaque's Moving Finger
-
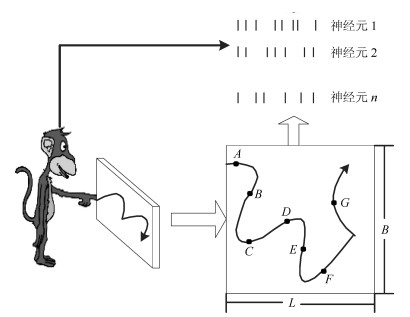
摘要: 利用猕猴运动皮层神经元峰电位数信号估计其手指移动位置是一神经解码问题, 通常采用时不变线性模型(Time-invariant linear model, TILM)来解决.本文分析了传统TILM模型的时间相关性问题, 依据猕猴手指移动位置的连续性特点, 采用一种新的模型去解码其手指移动位置, 称之为卷积空间模型(Convolution space model, CSM).与传统的模型相比, 卷积空间模型不但将当前时刻的状态与前一个时刻建立了相关, 而且与前多个时刻的状态也有相关.在实验中, 利用公开数据来评判本文方法的解码性能, 实验结果表明, 传统方法的解码误差要大于CSM模型的方法, 因此CSM模型具有更好的解码准确性.Abstract: It is a neural decoding problem to estimate the position of a macaque$'$s moving finger through neuron spike signals in motor cortex, which is usually solved by a time-invariant linear model (TILM). This paper analyzes the temporal correlation of the traditional TILM model. According to the continuity characteristics of the position of a macaque's moving finger, a new model is adopted to decode the finger movement, which is called CSM (Convolution space model). Compared with traditional decoding models, the CSM model can express that a state at the current time will be related to states at multiple previous times, rather than only one previous time. In experiments, we use the public data to evaluate the decoding performance of our method. The experimental results show that the CSM model has lower decoding errors than traditional methods and thus has better decoding accuracy.
-
Key words:
- Neural decoding /
- macaque's moving finger /
- CSM model /
- TILM model
1) 本文责任编委 秦涛 -
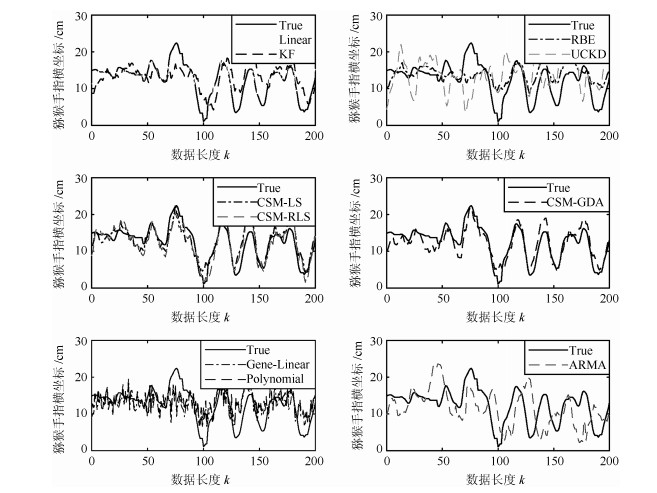
图 4 实验1中位置估计与手指移动真实位置曲线
Fig. 4 Position estimation and finger movement real position curve in experiment 1
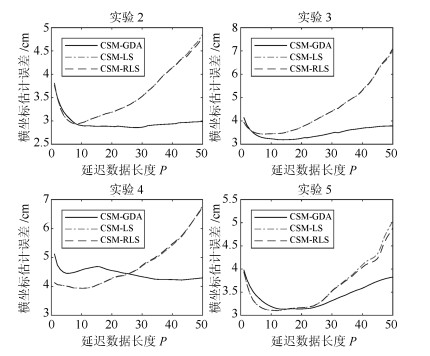
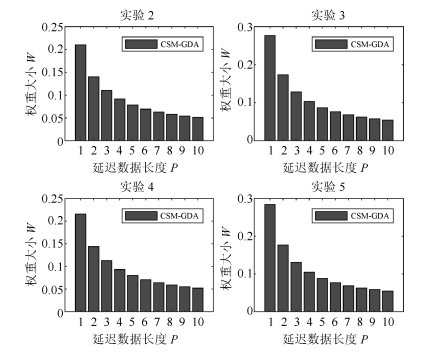
图 5 手指移动横坐标估计误差随延迟数据长度$P$的变化
Fig. 5 Finger movement abscissa estimation error with delay data length $P$ changes
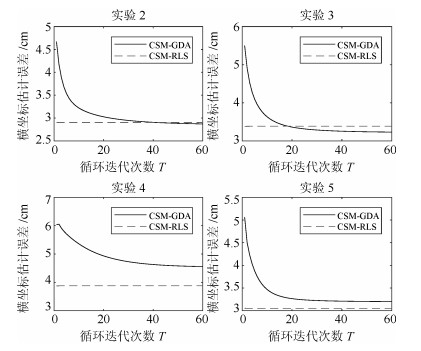
图 6 手指移动横坐标估计误差随迭代次数$T$的变化
Fig. 6 Finger movement abscissa estimation error with the number of iterations cycle $T$ changes
表 1 参数$P$对算法的训练复杂度
Table 1 Training complexity of parameter $P$ for algorithm
CSM模型算法 LS RLS GDA 复杂度 ${\rm O}(P^3)$ ${\rm O}(TP^3)$ ${\rm O}(TP^2)$  下载: 导出CSV
下载: 导出CSV
表 2 $X$轴和$Y$轴(括号内)的估计误差(cm) (保留三位)
Table 2 $X$-axis and $Y$-axis (in parentheses) estimated error (cm) (three places reserved)
算法$X(Y)$ 实验2 实验3 实验4 实验5 平均 Linear 3.813 (2.003) 3.941 (2.513) 4.135 (2.991) 3.919 (2.120) 3.952 (2.407) KF 3.060 (1.498) 3.908 (2.042) 4.540 (2.939) 3.637 (1.656) 3.786 (2.034) RBE 3.637 (1.727) 4.699 (2.295) 3.907 (2.400) 3.913 (1.936) 4.015 (2.089) UCKD 6.596 (4.235) 6.949 (5.504) 7.500 (5.775) 6.220 (4.868) 6.816 (5.096) CSM-LS 2.959 (1.411) 3.450 (2.154) 3.937 (2.921) 3.109 (1.724) 3.364 (2.052) CSM-RLS 2.964 (1.411) 3.443 (2.153) 3.957 (2.891) 3.106 (1.726) 3.368 (2.045) CSM-GDA 2.896 (1.500) 3.270 (2.095) 4.581 (2.406) 3.233 (1.709) 3.495 (1.927)
下载: 导出CSV
表 3 二维平面的估计误差(cm) (保留三位)
Table 3 The estimated error of the two-dimensional plane (cm) (three places reserved)
算法$XY$ 实验2 实验3 实验4 实验5 平均 Linear 4.307 4.674 5.103 4.455 4.635 KF 3.407 4.409 5.408 3.996 4.305 RBE 4.026 5.230 4.585 4.366 4.552 UCKD 7.839 8.865 9.466 7.896 8.787 CSM-LS 3.278 4.068 4.902 3.555 3.951 CSM-RLS 3.283 4.061 4.901 3.553 3.950 CSM-GDA 3.261 3.883 5.174 3.656 3.994
下载: 导出CSV
表 4 CSM模型算法的训练时间(保留三位)
Table 4 The training time of CSM model algorithm (three places reserved)
CSM模型算法 LS RLS GDA 训练时间(s) 1.061 23.853 1.285
下载: 导出CSV
-
[1] Campbell J, Sharma A. Visual cross-modal re-organization in children with cochlear implants. PLoS One, 2016, 11(1): e0147793 doi: 10.1371/journal.pone.0147793 [2] Wang W, Collinger J L, Degenhart A D, Tyler-Kabara E C, Schwartz A B, Moran D W, et al. An electrocorticographic brain interface in an individual with tetraplegia. PLoS One, 2013, 8(2): e55344 doi: 10.1371/journal.pone.0055344 [3] 李远清.脑机接口技术在意识障碍领域应用的前景展望.中华神经创伤外科电子杂志, 2015, 1(2): 60-61Li Yuan-Qing. Application prospect of brain computer interface technology in the field of consciousness disorders. Chinese Journal of Neurotraumatic Surgery (Electronic Edition), 2015, 1(2): 60-61 [4] Wang Y W, Wang F, Xu K, Zhang Q S, Zhang S M, Zheng X X. Neural control of a tracking task via attention-gated reinforcement learning for brain-machine interfaces. IEEE Transactions on Neural Systems and Rehabilitation Engineering, 2015, 23(5): 458-467 [5] 郑筱祥, 王怡雯, 张韶岷, 张巧生.猴子PMd区脑电解码抓握手势及机械手实时控制.科技创新导报, 2016, 13(12): 167-168 doi: 10.3969/j.issn.1674-098X.2016.12.093Zheng Xiao-Xiang, Wang Yi-Wen, Zhang Shao-Min, Zhang Qiao-Sheng. Decoding grasp movement from monkey premotor cortex for real-time prothetic hand control. Science and Technology Innovation Herald, 2016, 13(12): 167-168 doi: 10.3969/j.issn.1674-098X.2016.12.093 [6] Bouton C E, Shaikhouni A, Annetta N V, Bockbrader M A, Friedenberg D A, Nielson D M, et al. Restoring cortical control of functional movement in a human with quadriplegia. Nature, 2016, 533(7602): 247-250 doi: 10.1038/nature17435 [7] 侯增广, 赵新刚, 程龙, 王启宁, 王卫群.康复机器人与智能辅助系统的研究进展.自动化学报, 2016, 42(12): 1765-1779 doi: 10.16383/j.aas.2016.y000006Hou Zeng-Guang, Zhao Xin-Gang, Cheng Long, Wang Qi-Ning, Wang Wei-Qun. Recent advances in rehabilitation robots and intelligent assistance systems. Acta Automatica Sinica, 2016, 42(12): 1765-1779 doi: 10.16383/j.aas.2016.y000006 [8] Wallisch P, Lusignan M E, Benayoun M D, Baker T L, Dickey A S, Hatsopoulos N G. MATLAB for Neuroscientists (Second edition). London: Elsevier, 2014. [9] Kass R E, Eden U T, Brown E N. Analysis of Neural Data. New York: Springer, 2014. [10] Hatsopoulos N G, Ojakangas C L, Paninski L, Donoghue J P. Information about movement direction obtained from synchronous activity of motor cortical neurons. Proceedings of the National Academy of Sciences of the United States of America, 1998, 95(26): 15706-15711 doi: 10.1073/pnas.95.26.15706 [11] Hatsopoulos N G, Xu Q Q, Amit Y. Encoding of movement fragments in the motor cortex. The Journal of Neuroscience, 2007, 27(19): 5105-5114 doi: 10.1523/JNEUROSCI.3570-06.2007 [12] Moran D W, Schwartz A B. Motor cortical representation of speed and direction during reaching. Journal of Neurophysiology, 1999, 82(5): 2676-2692 doi: 10.1152/jn.1999.82.5.2676 [13] Reynaud-Bouret P, Rivoirard V, Grammont F, Tuleau-Malot C. Goodness-of-fit tests and nonparametric adaptive estimation for spike train analysis. Journal of Mathematical Neuroscience, 2014, 4: 3 doi: 10.1186/2190-8567-4-3 [14] Georgopoulos A P, Kettner R E, Schwartz A B. Primate motor cortex and free arm movements to visual targets in three-dimensional space. Ⅱ. Coding of the direction of movement by a neuronal population. Journal of Neurophysiology, 1988, 8(8): 2928-2937 [15] Georgopoulos A P, Lurito J T, Petrides M, Schwartz A B, Massey J T. Mental rotation of the neuronal population vector. Science, 1989, 243(4888): 234-236 doi: 10.1126/science.2911737 [16] Vargas-Irwin C E, Shakhnarovich G, Yadollahpour P, Mislow J M K, Black M J, Donoghue J P. Decoding complete reach and grasp actions from local primary motor cortex population. Journal of Neuroscience, 2010, 30(29): 9659-9669 doi: 10.1523/JNEUROSCI.5443-09.2010 [17] O'Doherty J E, Lebedev M A, Ifft P J, Zhuang K Z, Shokur S, Bleuler H, et al. Active tactile exploration enabled by a brain-machine-brain interface. Nature, 2011, 479(7372): 228-231 doi: 10.1038/nature10489 [18] Serruya M D, Hatsopoulos N G, Paninski L, Fellows M R, Donoghue J P. Instant neural control of a movement signal. Nature, 2002, 416(6877): 141-142 doi: 10.1038/416141a [19] Warland D K, Reinagel P, Meister M. Decoding visual information from a population of retinal ganglion cells. Journal of Neurophysiology, 1997, 78(5): 2336-2350 doi: 10.1152/jn.1997.78.5.2336 [20] Velliste M, Perel S, Spalding M C, Whitford A S, Schwartz A B. Cortical control of a prosthetic arm for self-feeding. Nature, 2008, 453(7198): 1098-1101 doi: 10.1038/nature06996 [21] Shanechi M M, Wornell G W, Williams Z M, Brown E N. Feedback-controlled parallel point process filter for estimation of goal-directed movements from neural signals. IEEE Transactions on Neural Systems and rehabilitation Engineering, 2013, 21(1): 129-140 doi: 10.1109/TNSRE.2012.2221743 [22] Chang Y H, Chen M, Shanechi M, Carmena J M, Tomlin C. A design of neural decoder by reducing discrepancy between manual control (MC) and brain control (BC). In: Proceedings of the 2014 European Control Conference. Strasbourg, France: IEEE, 2014. 516-521 [23] 薛明龙, 吴海锋, 曾玉.无监督的猕猴运动皮层锋电位信号CKF解码.自动化学报, 2017, 43(2): 302-312 doi: 10.16383/j.aas.2017.c160065Xue Ming-Long, Wu Hai-Feng, Zeng Yu. Unsupervised CKF decoding for macaque motor cortical spikes. Acta Automatica Sinica, 2017, 43(2): 302-312 doi: 10.16383/j.aas.2017.c160065 [24] Xue M L, Wu H F, Zeng Y, Yang K. Estimate a macaque's finger trajectory using unsupervised cubature Kalman filtering decoding. In: Proceedings of the 2016 IEEE Advanced Information Management, Communicates, Electronic and Automation Control Conference (IMCEC). Xi'an, China: IEEE, 2016. 605-609 [25] Hotson G, Smith R J, Rouse A G, Schieber M H, Thakor N V, Wester B A. High precision neural decoding of complex movement trajectories using recursive Bayesian estimation with dynamic movement primitives. IEEE Robotics and Automation Letters, 2016, 1(2): 676-683 doi: 10.1109/LRA.2016.2516590 [26] 李宏宝.猕猴手臂避障规划与执行过程中背侧运动前区皮层的表征与解码[博士学位论文], 浙江大学, 中国, 2017Li Hong-Bao. PMD representation and decoding of monkey reach plan and execution during obstacle avoidance task[Ph. D. dissertation], Zhejiang University, China, 2017 [27] 张毅, 尹春林, 蔡军, 罗久飞. Bagging RCSP脑电特征提取算法.自动化学报, 2017, 43(11): 2044-2050 doi: 10.16383/j.aas.2017.c160094Zhang Yi, Yin Chun-Lin, Cai Jun, Luo Jiu-Fei. Bagging RCSP algorithm for extracting EEG feature. Acta Automatica Sinica, 2017, 43(11): 2044-2050 doi: 10.16383/j.aas.2017.c160094 [28] Brockwell A E, Rojas A L, Kass R E. Recursive Bayesian decoding of motor cortical signals by particle filtering. Journal of Neurophysiology, 2004, 91(4): 1899-1907 doi: 10.1152/jn.00438.2003 [29] Wu W, Shaikhouni A, Donoghue J P, Black M J. Closed-loop neural control of cursor motion using a Kalman filter. In: Proceedings of the 26th Annual International Conference of the IEEE Engineering in Medicine and Biology Society (EMBC). San Francisco, CA, USA: IEEE, 2004. 4126-4129 [30] Wu W, Gao Y, Bienenstock E, Donoghue J P, Black M J. Bayesian population decoding of motor cortical activity using a Kalman filter. Neural Computation, 2006, 18(1): 80-118 doi: 10.1162/089976606774841585 [31] Haykin S. Neural Networks and Learning Machines (Third edition). Upper Saddle River, NJ: Prentice Hall, 2008. [32] 张贤达.现代信号处理.第3版.北京:清华大学出版社, 2015.Zhang Xian-Da. Modern Signal Processing (Third edition). Beijing: Tsinghua University Press, 2015. [33] Raschka S, Mirjalili V. Python Machine Learning (Second edition). Birmingham, UK: Packt Publishing, 2017. -

 下载:
下载:

计量
- 文章访问数: 1010
- HTML全文浏览量: 324
- PDF下载量: 131
- 被引次数: 0
