

-
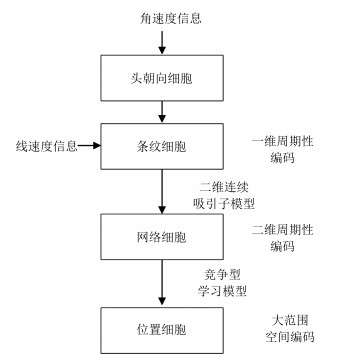
摘要: 海马结构空间细胞的放电活动被认为能够形成对环境内在地图的表达,即所谓的认知地图.先前的仿生环境认知地图构建方法(例如RatSLAM)以及传统的SLAM方法均缺乏足够的生理学依据,不能准确地体现出生物在导航中的生理学现象和认知功能实现过程.本文模仿海马结构空间细胞的认知机理提出了一种构建精确的环境认知地图的方法,其特点在于通过构建统一的空间细胞吸引子计算模型对自运动线索进行路径积分;网格细胞和位置细胞对环境的表达来源于条纹细胞的前向驱动作用;通过环境的颜色深度图像进行闭环检测,对空间细胞路径积分进行误差修正,最终生成精确的环境认知地图.该认知地图是一种拓扑度量地图,包含了环境特征点坐标、视觉线索以及特定位点的拓扑关系.本文通过仿真实验和机器人平台物理实验验证了方法的有效性,研究成果为仿海马认知机理的机器人导航方法研究奠定了基础.Abstract: Spatial cells in hippocampus play a functional role in representation and processing of spatial information, which appear to provide a basis for cognitive map:a representation of environment. Most prior biomimetic map building algorithms, such as RatSLAM algorithm or traditional SLAM methods, have little biological fidelity to the hippocampal formation. In this paper, a neural network model based on the behavioral and neurophysiological mechanisms of the spatial cells is constructed, and is applied to building the accurate cognitive map of real environments. The proposed algorithm has a uniform calculation method for spatial cells based on continuous attractor network dynamics to integrate self-motion cues, which can reproduce grid cells firing responses and place cells firing fields via feedforward inputs from band cells. RGB-D images serve as visual cues for loop closure detection and correcting the accumulative errors intrinsically associated with the path integration mechanism, which contributes to building spatial cognitive maps of indoor environments on a mobile robot. A cognitive map is a fine-grained topological-metric map. A node in the cognitive map is constructed by associating the major peak of place cell population activities with corresponding visual cues and the transition stores the change in positions. Simulation experiments and physical experiments with a mobile robot have verified the effectiveness of the algorithm. The proposed algorithm may provide a foundation for robotic navigation.
-
Key words:
- Cognitive map /
- hippocampal formation /
- spatial cells /
- band cells /
- loop closure detection
1) 本文责任编委 徐德 -

图 8 位置细胞的连续吸引子模型表达
Fig. 8 The expression of the continuous attractor model of place cells
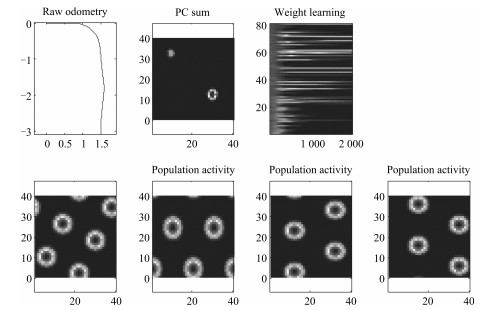
图 13 老鼠移动轨迹上网格细胞和位置细胞放电率图
Fig. 13 The firing rate of place cells and grid cells based on mouse trajectory

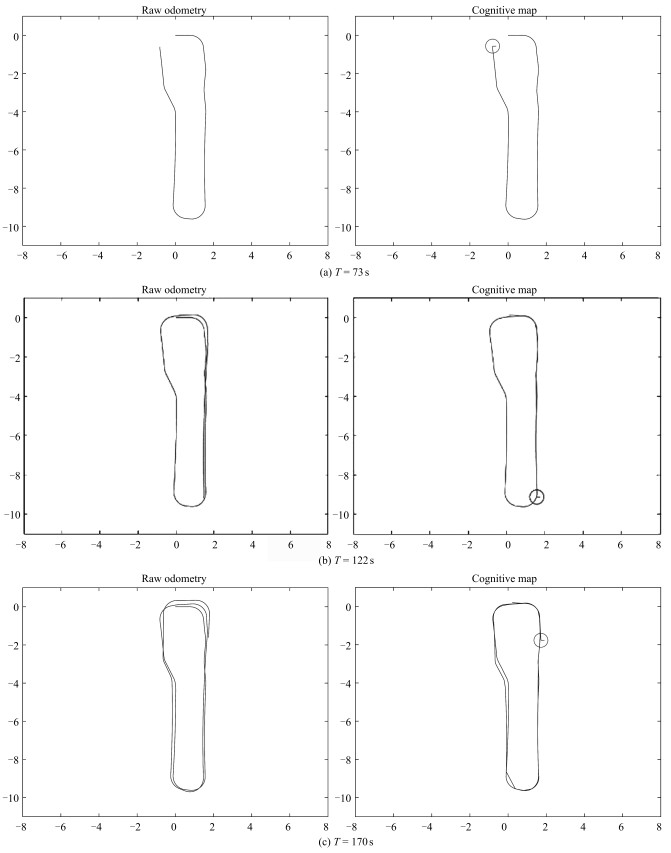
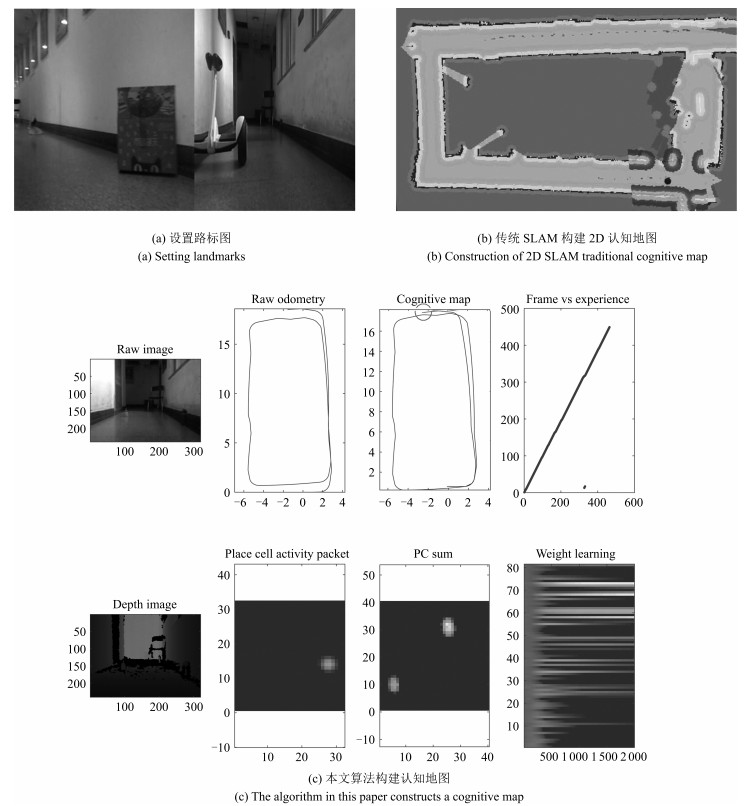
图 17 闭环检测与空间细胞放电重置过程
Fig. 17 Closed loop detection and space cell discharge reset process

图 18 位置细胞相互竞争前后的放电率图
Fig. 18 The firing rate before and after the place cell competition

图 23 网格间距为4 cm时单个神经元的响应图
Fig. 23 The response of a single neuron when the grid space is 4 cm
表 1 参数设置
参数 值 $n_X=n_Y$ 32 $k_p$ 7 $\varphi$ 0.00002 $\rho$ 20 $c_t$ 1 $S_{th}$ 1 $\vartheta$ 0.5 $\mu_R$ 0.65 $\mu_D$ 0.35  下载: 导出CSV
下载: 导出CSV
-
[1] Tolman E C. Purposive Behavior in Animals and Men. New York, USA:University of California Press, 1932. [2] McNaughton B L, Battaglia F P, Jensen O, Moser E I, Moser M B. Path integration and the neural basis of the "cognitive map". Nature Reviews Neuroscience, 2006, 7(8):663-678 doi: 10.1038/nrn1932 [3] Steele R J, Morris R G. Delay-dependent impairment of a matching-to-place task with chronic and intrahippocampal infusion of the NMDA-antagonist D-AP5. Hippocampus, 1999, 9(2):118-136 doi: 10.1002/(ISSN)1098-1063 [4] Steffenach H A, Witter M, Moser M B, Moser E I. Spatial memory in the rat requires the dorsolateral band of the entorhinal cortex. Neuron, 2005, 45(2):301-313 doi: 10.1016/j.neuron.2004.12.044 [5] Taube J S, Kesslak J P, Cotman C W. Lesions of the rat postsubiculum impair performance on spatial tasks. Behavioral and Neural Biology, 1992, 57(2):131-143 doi: 10.1016/0163-1047(92)90629-I [6] O'Keefe J, Dostrovsky J. The hippocampus as a spatial map. Preliminary evidence from unit activity in the freely-moving rat. Brain Research, 1971, 34(1):171-175 doi: 10.1016/0006-8993(71)90358-1 [7] O'Keefe J. Place units in the hippocampus of the freely moving rat. Experimental Neurology, 1976, 51(1):78-109 doi: 10.1016/0014-4886(76)90055-8 [8] Tsao A, Moser M B, Moser E I. Traces of experience in the lateral entorhinal cortex. Current Biology, 2013, 23(5):399-405 doi: 10.1016/j.cub.2013.01.036 [9] Taube J S, Muller R U, Ranck Jr J B. Head-direction cells recorded from the postsubiculum in freely moving rats. Ⅱ. Effects of environmental manipulations. The Journal of Neuroscience, 1990, 10(2):436-447 http://www.jneurosci.org/content/jneuro/10/2/436.full.pdf [10] Sharp P E. Multiple spatial/behavioral correlates for cells in the rat postsubiculum:multiple regression analysis and comparison to other hippocampal areas. Cerebral Cortex, 1996, 6(2):238-259 doi: 10.1093/cercor/6.2.238 [11] O'Keefe J, Burgess N. Geometric determinants of the place fields of hippocampal neurons. Nature, 1996, 381(6581):425-428 doi: 10.1038/381425a0 [12] Mittelstaedt M L, Mittelstaedt H. Homing by path integration in a mammal. Naturwissenschaften, 1980, 67(11):566-567(in German) doi: 10.1007/BF00450672 [13] Hafting T, Fyhn M, Molden S, Moser M B, Moser E I. Microstructure of a spatial map in the entorhinal cortex. Nature, 2005, 436(7052):801-806 doi: 10.1038/nature03721 [14] Solstad T, Moser E I, Einevoll G T. From grid cells to place cells:a mathematical model. Hippocampus, 2006, 16(12):1026-1031 doi: 10.1002/hipo.v16:12 [15] Moser E I, Roudi Y, Witter M P, Kentros C, Bonhoeffer T, Moser M B. Grid cells and cortical representation. Nature Reviews Neuroscience, 2014, 15(7):466-481 doi: 10.1038/nrn3766 [16] Burgess N, Barry C, O'Keefe J. An oscillatory interference model of grid cell firing. Hippocampus, 2007, 17(9):801-812 doi: 10.1002/(ISSN)1098-1063 [17] Hasselmo M E, Giocomo L M, Zilli E A. Grid cell firing may arise from interference of theta frequency membrane potential oscillations in single neurons. Hippocampus, 2007, 17(12):1252-1271 doi: 10.1002/(ISSN)1098-1063 [18] Si B, Romani S, Tsodyks M. Continuous attractor network model for conjunctive position-by-velocity tuning of grid cells. PLoS Computational Biology, 2014, 10(4):e1003558 doi: 10.1371/journal.pcbi.1003558 [19] Erdem U M, Hasselmo M. A goal-directed spatial navigation model using forward trajectory planning based on grid cells. European Journal of Neuroscience, 2012, 35(6):916-931 doi: 10.1111/ejn.2012.35.issue-6 [20] Fuhs M C, Touretzky D S. A spin glass model of path integration in rat medial entorhinal cortex. The Journal of Neuroscience, 2006, 26(16):4266-4276 doi: 10.1523/JNEUROSCI.4353-05.2006 [21] Burak Y, Fiete I R. Accurate path integration in continuous attractor network models of grid cells. PLoS Computational Biology, 2009, 5(2):e1000291 doi: 10.1371/journal.pcbi.1000291 [22] Krupic J, Burgess N, O'Keefe J. Neural representations of location composed of spatially periodic bands. Science, 2012, 337(6096):853-857 doi: 10.1126/science.1222403 [23] Raudies F, Hasselmo M E. Differences in visual-spatial input may underlie different compression properties of firing fields for grid cell modules in medial entorhinal cortex. PLoS Computational Biology, 2015, 11(11):e1004596 doi: 10.1371/journal.pcbi.1004596 [24] Yuan M L, Tian B, Shim V A, Tang H, Li H. An entorhinal-hippocampal model for simultaneous cognitive map building. In:Proceedings of the 29th AAAI Conference on Artificial Intelligence. Austin, TX:AAAI, 2015. [25] Grossberg S, Pilly P K. How entorhinal grid cells may learn multiple spatial scales from a dorsoventral gradient of cell response rates in a self-organizing map. PLoS Computational Biology, 2012, 8(10):e1002648 doi: 10.1371/journal.pcbi.1002648 [26] Pilly P K, Grossberg S. Spiking neurons in a hierarchical self-organizing map model can learn to develop spatial and temporal properties of entorhinal grid cells and hippocampal place cells. PLoS One, 2013, 8(4):e60599 doi: 10.1371/journal.pone.0060599 [27] Cuperlier N, Quoy M, Gaussier P. Neurobiologically inspired mobile robot navigation and planning. Frontiers in Neurorobotics, 2007, 1:Article No. 3 [28] Jauffret A, Cuperlier N, Gaussier P. Multimodal integration of visual place cells and grid cells for robots navigation. In:Proceedings of the 2nd Symposium on Biology of Decision Making. Paris, France:BDM, 2012. 136-145 [29] Milford M J, Wyeth G F, Prasser D. RatSLAM:a hippocampal model for simultaneous localization and mapping. In:Proceedings of the 2004 IEEE International Conference on Robotics and Automation. New Orleans, LA, USA:IEEE, 2004. 403-408 [30] Milford M, Wyeth G, Prasser D. RatSLAM on the edge:revealing a coherent representation from an overloaded rat brain. In:Proceedings of the 2006 IEEE/RSJ International Conference on Intelligent Robots and Systems. Beijing, China:IEEE, 2006. 4060-4065 [31] Milford M J, Wyeth G F. Mapping a suburb with a single camera using a biologically inspired SLAM system. IEEE Transactions on Robotics, 2008, 24(5):1038-1053 doi: 10.1109/TRO.2008.2004520 [32] Milford M J, Wyeth G F. Single camera vision-only SLAM on a suburban road network. In:Proceedings of the 2008 IEEE International Conference on Robotics and Automation. Pasadena, CA, USA:IEEE, 2008. 3684-3689 [33] Erdem U M, Hasselmo M E. A biologically inspired hierarchical goal directed navigation model. Journal of Physiology, Paris, 2014, 108(1):28-37 doi: 10.1016/j.jphysparis.2013.07.002 [34] 王楠, 马书根, 李斌, 王明辉, 赵明扬.震后建筑内部层次化SLAM的地图模型转换方法.自动化学报, 2015, 41(10):1723-1733 http://www.aas.net.cn/CN/Y2015/V41/I10/1723Wang Nan, Ma Shu-Gen, Li Bin, Wang Ming-Hui, Zhao Ming-Yang. A model transformation of map representation for hierarchical SLAM that can be used for after-earthquake buildings. Acta Automatica Sinica, 2015, 41(10):1723-1733 http://www.aas.net.cn/CN/Y2015/V41/I10/1723 [35] 肖云涛, 欧林林, 俞立.基于线性时序逻辑的最优巡回路径规划.自动化学报, 2014, 40(10):2126-2133 http://www.aas.net.cn/CN/Y2014/V40/I10/2126Xiao Yun-Tao, Ou Lin-Lin, Yu Li. Optimal patrolling path planning via linear temporal logic. Acta Automatica Sinica, 2014, 40(10):2126-2133 http://www.aas.net.cn/CN/Y2014/V40/I10/2126 [36] 徐博, 白金磊, 郝燕玲, 高伟, 刘亚龙.多AUV协同导航问题的研究现状与进展.自动化学报, 2015, 41(3):445-461 http://www.aas.net.cn/CN/Y2015/V41/I3/445Xu Bo, Bai Jin-Lei, Hao Yan-Ling, Gao Wei, Liu Ya-Long. The research status and progress of cooperative navigation for multiple AUVs. Acta Automatica Sinica, 2015, 41(3):445-461 http://www.aas.net.cn/CN/Y2015/V41/I3/445 [37] 陈成, 何玉庆, 卜春光, 韩建达.基于四阶贝塞尔曲线的无人车可行轨迹规划.自动化学报, 2015, 41(3):486-496 doi: 10.16383/j.aas.2015.c140295Chen Cheng, He Yu-Qing, Bu Chun-Guang, Han Jian-Da. Feasible trajectory generation for autonomous vehicles based on quartic bézier curve. Acta Automatica Sinica, 2015, 41(3):486-496 doi: 10.16383/j.aas.2015.c140295 [38] Ranck J B. Head direction cells in the deep cell layer of dorsolateral presubiculum in freely moving rats. Electrical Activity of the Archicortex, Budapest:Akadémiai Kiadó, 1984. [39] Raudies F, Chapman W, Brandon P, Hasselmo M E. Head direction is coded more strongly than movement direction in a population of entorhinal neurons. Brain Research, 2015, 1621:355-367 doi: 10.1016/j.brainres.2014.10.053 [40] Mizumori S J, Williams J D. Directionally selective mnemonic properties of neurons in the lateral dorsal nucleus of the thalamus of rats. The Journal of Neuroscience, 1993, 13(9):4015-4028 http://citeseerx.ist.psu.edu/showciting?cid=3849877 [41] Taube J S, Muller R U, Ranck Jr J B. Head-direction cells recorded from the postsubiculum in freely moving rats. I. Description and quantitative analysis. The Journal of Neuroscience, 1990, 10(2):420-435 http://citeseerx.ist.psu.edu/viewdoc/summary?doi=10.1.1.328.9864 [42] Knierim J J, Kudrimoti H S, McNaughton B L. Place cells, head direction cells, and the learning of landmark stability. The Journal of Neuroscience, 1995, 15(3 Pt 1):1648-1659 https://jhu.pure.elsevier.com/en/publications/place-cells-head-direction-cells-and-the-learning-of-landmark-sta-4 [43] Kropff E, Carmichael J E, Moser M B, Moser E I. Speed cells in the medial entorhinal cortex. Nature, 2015, 523(7561):419-424 doi: 10.1038/nature14622 [44] Juavinett A L, Callaway E M. Pattern and component motion responses in mouse visual cortical areas. Current Biology, 2015, 25(13):1759-1764 doi: 10.1016/j.cub.2015.05.028 [45] Couey J J, Witoelar A, Zhang S J, Zheng K, Ye J, Dunn B, Czajkowski R, Moser M B, Moser E, Roudi Y, Witter M P. Recurrent inhibitory circuitry as a mechanism for grid formation. Nature Neuroscience, 2013, 16:318-324 doi: 10.1038/nn.3310 [46] Si B L, Treves A. The role of competitive learning in the generation of dg fields from ec inputs. Cognitive Neurodynamics, 2009, 3(2):177-187 doi: 10.1007/s11571-009-9079-z [47] 于乃功, 王琳, 李倜, 陈焕朝.网格细胞到位置细胞的竞争型神经网络模型.控制与决策, 2015, 30(8):1372-1378 http://www.intsci.ac.cn/ai/nn.htmlYu Nai-Gong, Wang Lin, Li Ti, Chen Huan-Zhao. Competitive neural network model from grid cells to place cells. Control and Decision, 2015, 30(8):1372-1378 http://www.intsci.ac.cn/ai/nn.html [48] Bonnevie T, Dunn B, Fyhn M, Hafting T, Derdikman D, Kubie J L, Roudi Y, Moser E I, Moser M B. Grid cells require excitatory drive from the hippocampus. Nature Neuroscience, 2013, 16(3):309-317 doi: 10.1038/nn.3311 [49] Winter S S, Mehlman M L, Clark B J, Taube J S. Passive transport disrupts grid signals in the parahippocampal cortex. Current Biology, 2015, 25(19):2493-2502 doi: 10.1016/j.cub.2015.08.034 [50] Tian B, Shim V A, Yuan M L, Srinivasan C, Tang H J, Li H Z. RGB-D based cognitive map building and navigation. In:Proceedings of the 2013 IEEE/RSJ International Conference on Intelligent Robots and Systems (IROS). Tokyo, Japan:IEEE, 2013. 1562-1567 [51] 潘超, 刘建国, 李峻林.昆虫视觉启发的光流复合导航方法.自动化学报, 2015, 41(6):1102-1112 http://www.aas.net.cn/CN/Y2015/V41/I6/1102Pan Chao, Liu Jian-Guo, Li Jun-Lin. An optical flow-based composite navigation method inspired by insect vision. Acta Automatica Sinica, 2015, 41(6):1102-1112 http://www.aas.net.cn/CN/Y2015/V41/I6/1102 -

 下载:
下载:













计量
- 文章访问数: 2932
- HTML全文浏览量: 695
- PDF下载量: 801
- 被引次数: 0
